Дьявол таится в деталях. Парадоксально, но факт!
 Отношения на парадоксах естественных технологий
Отношения на парадоксах естественных технологий
Еще на заре развития кибернетики, F.Grodins в своей книге "Control theory and biological systems" (Columbia Univ. Proc. — London, 1963) писал: "...физические проблемы обычно настолько просты, что всегда легко обнаружить наличие парадокса в интерпретации того или иного явления, тогда как биологические проблемы обычно настолько сложны, что никогда нет полной уверенности в наличии или отсутствии такого парадокса". Навряд ли такое противопоставление можно считать справедливым, однако наше исследование не стало исключением.
Парадоксы, как логические противоречия, из которых казалось бы, невозможно найти выхода, были известны еще античным мыслителям. С того времени осталось множество нерешенных парадоксов ("Куча", "Лжец", "Ахиллес и черепаха и др."), доставивших много труда и древнегреческим и современным математикам, логикам и философам, пытавшихся различными методами преодолеть возникшие противоречия. Появилось множество парадоксов, поставивших под сомнение истинность теории множеств, как в неполной мере уяснившей собственные функциональные закономерности и логические основания.
В статье " Феномен физического времени ", мы попытались установить понятие "живое", как единство потоков времени, клеточного транспорта и дискрет информации, определенных на глубинных отношениях "пространство-время" ("структура-функция"). Такое единство реализуется системой локального гомеостазиса как биологической естественно-технологической системой программного управления. Обладая свойствами гомеостата, она стабилизирует, относительно заданного уровня в потоке времени, процесс поддержания транспортных потоков множеств озлокачествляемых клеток, образуя систему гомеореза — наиболее сложную самоуправляемую и самодостаточную биологическую систему управления потоками. Понимание механизмов гомеореза, даже с учетом уже полученных знаний, недостаточно для ее четкой интерпретации в количественных терминах.
Легче всего объяснить трудность математического изучения деталей этой проблемы сложностью биологических систем, объективно обуславливающей вероятность порождения парадоксов. Но парадоксы могут появляться и в области истинных суждений: парадоксально, что земля движется вокруг солнца, что вода состоит из двух легко воспламеняющихся газов. Поэтому появление парадоксов само по себе не удивительно: на клеточных множествах удивительны предмет парадокса, как порожденный объект мысли и обескураживающие чувства внутреннего состояния, возникающие из обсуждения дилемм, порождаемых этим объектом.
Проблема пространственно-временных отношений — одна из сложнейших и запутанных проблем как физики, так и биологии и неудивительно, что при изучении этой проблемы на конкретной предметной области, мы столкнулись с совершенно неожиданным, странным и озадачивающим предметом парадокса. Предмет возникшего парадокса породил проблему оценки отношения изначального соответствия внешнего проявления, "физического" образа опухоли и ее математического отображения. Оказалось, что обобщающее название злокачественной опухоли "рак", которым наделили ее врачи средневековья, отражается в абстрактном пространстве характеристической пирамиды графом агрессивности локального гомеостазиса, покроенным формальным путем на мере озлокачествления. Объяснения существенности предмета парадокса, источника появления и значимости в рамках языка изучаемого мира отношений не найдено. Попытки дать ему реалистичное объяснение приводят в тупик или граничат с мистикой, а интерпретации выходят за пределы логики исследования, образуя отношения внешнего, относительно рассматриваемого мира, порядка.
Парадоксальность ситуации удивляет тем обстоятельством, что в процессе исследований, неожиданно проявивший себя на графе агрессивности для канонического состояния в пространстве характеристической пирамиды эффект, никак не предугадывался, в логику и последовательность анализа не закладывался, и априори фактически воспроизведен быть не может. Этот граф, отражая единство на потоках клеточных множеств, квантов времени и дискрет информации, стал обозримым в результате применения последовательно-логического анализа закономерностей ракового гомеостата обычными для общей теории систем методами, результаты использования которых обозримы на предыдущих публикациях.
Сетевое пространство и изолированность клеточных сетей.
Строя кривые частотных распределений, мы определяли частости появления каждого i-го подмножества клеток, обладающих тем или иным значением xi ядерно-цитоплазматического отношения, группируя в них такие клетки, измерения по которым были достаточно близки на интервале некоторого значения xi, зависящего от погрешностей измерений и обработки, в том числе от округления результата вычислений. Поэтому, говоря о клеточных денотатах констант состояния, как и о всех других множествах, отраженных значениями обобщенного показателя озлокачествления на вершинах графа, мы рассматриваем наблюденные множества, как ограниченные сверху и снизу множества метрического пространства локального гомеостазиса, граница которого конечна и выражена на мере Гомпертца.
Каждое такое множество на вершинах графа можно рассматривать в качестве ядра некоторого множества M, образующегося по построению объединением множества клеток, обладающих наблюденными в клинике аргументами параметра N<I,j> обобщенного показателя озлокачествления и подмножеств клеток его ближайших окрестностей некоторого диаметра r. Диаметр r окрестностей этого множества определяется степенью конкретизации и точностью обработки клинических данных исходного описания. Его величина зависит от максимальных погрешностей оценки аргументов x,L обобщенного показателя N<I,j>, но в большей степени — от грубости оценки показателя прозрачности ядер L, превосходящей погрешности оценки ядерно-цитоплазматических отношений. Поэтому значение r определяется погрешностями, сопровождающими оценку прозрачности ядер. В принципе же из за неравноточности данных исходного описания, ограничения множеств метрического пространства локального гомеостазиса характеризуются эллипсоидом вращения, полуоси которого соответствуют погрешностям оценок аргументов x,L показателя озлокачествления.
Множества клеток ближайших окрестностей выделить в самостоятельные "окрестностные" подмножества не представляется возможным, вследствие непреодолимых трудностей наблюдения текущих состояний биологического субстрата с одной стороны, и подготовки данных исходного описания с повышением уровня конкретизации, с другой. Такие окрестностные множества в любом случае могут только сокращать свою мощность, но сохраняются при любой степени конкретизации, отражая лишь изменения параметров окрестностей того множества клеток, которые образовали ядро. Поэтому отображение подмножеств клеток, образующих на множестве окрестностных ситуационных сетей каждой из вершин графа агрессии ("Организация топологического пространства локального гомеостазиса"), как и подмножеств клеток и их ситуационных сетей всех других графов, рассматривается как отображение ситуационной сети на модели клеточной ситуации ядер множеств, например — клеток денотата дисплазионной ядерно-цитоплазматической константы.
Ядро множества подмножеств клеток агрессии ракового гомеостата в модели, образовано объединением всего наблюденного множества клеток на ситуационных сетях ближайших окрестностей множества подмножеств баланса эквивалентности. Фактически же, в субстрате существуют ситуационные сети как на клеточных ситуациях ядра множеств, так и на подсетях, образующиеся на ситуациях множеств клеток его ближайших окрестностей. Эти подсети конкатенированы в единую ситуационную сеть Подсети окрестностей вершин графа однако, остаются не отображаемыми в моделях любой степени конкретизации.
Под ситуационной сетью, образуемой множеством функционально связанных клеточных подмножеств на вершине графа, понимается ориентированный ситуационный граф, изначально не содержащий замкнутых контуров. Ситуационные сети на множествах клеток функционально связанных подмножеств и множеств клеток-денотатов констант состояний изолированные. Говорят, что на таком графе имеется ровно одна вершина — вход сети, в которую не входит ни одна дуга, и ровно одна вершина — выход сети, из которой не исходит ни одна дуга. Дуги ситуационного графа отражают характер потоков информационно-управляющих отношений. Нарушение изолированности сетей множеств клеток-денотатов константы состояния гомеостазиса означает подготовку гомеостазического переключения. Нарушение изолированности сетей множеств клеток-денотатов констант состояний порождения, означает образование цепочки множеств агрессивных клеток-денотатов констант состояний развития и озлокачествления.
Естественно-технологическое гомеостатическое и многокомпонентное сохранение локального гомеостазиса, охватывая на шагах рекурсии управления все множество изменений текущих состояний, порождаемых множеством клеток баланса эквивалентности, реализуется прорывом изолированности клеточных управляющих отношений на ситуационных сетях. Говоря о графах, мы имеем в виду как множество межклеточных, на сетях подмножеств клеток вершин графа, так и групповых взаимодействий, на сетях обмена информацией клеточных множеств, образующих множество его вершин.
Прорывы изолированности управляющих отношений на сетях множеств клеток-денотатов состояний, приводят к перетоку множеств клеток баланса эквивалентности в множество клеток баланса толерантности и образуют направленность клеточного транспорта, отражаемую, с одной стороны траекторией изображающей точки в фазовом пространстве и с другой — пространственным графом на топологии клеточных множеств, четвертую мерность на котором, определяет Ньютоновское время наблюдения процесса.
Пространство клеточных коммуникаций графа агрессивности. Парадокс канцерогенеза.
Пространство клеточных коммуникаций графа агрессивности локального гомеостазиса определяется топографией его вершин и пространствами сетей множеств клеток-денотатов состояний. Границы этого пространства отображаются положением ядер множеств, отраженных в значения обобщенного показателя озлокачествления на вершинах графа агрессии и не учитывают пространств подсетей окрестностей, вошедших в ядро как в открытое множество.
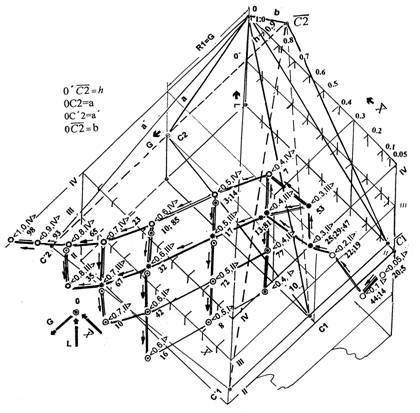
Предположим, мы хотим отобразить топографический контур пространства этих сетей. Для этого мы должны учесть максимальные погрешности +-L оценки градаций L прозрачности ядер в плоскостях их отсчета и аналогичные погрешности +-x оценки ядерно-цитоплазматического отношения x в соответствующих плоскостях. Учет численных оценок этих погрешностей, привел бы к построению множеств эллипсоидов на множествах пространств окрестностей вершин графа. Однако, в силу иллюстрационного характера описания, представляется допустимым очертить контур графа агрессии в пространстве характеристической пирамиды, ограничиваясь отклонением от положения его вершин на величину в пределах +-(8÷12)%, соответствующую максимальным погрешностям субъективной оценки градаций прозрачности ядер. На рисунке 1 приведен пространственный граф достижения канонического состояния, как граф агрессии порождения, развития и озлокачествления. Текущие значения атрибутов потоков транспортируемых множеств, отражаются идентификаторами, выраженными на обобщенном параметре озлокачествления. Поток времени рассматривается как его редукция. Этот граф отличается от рассмотренного ранее графа агрессии, множеством вершин и отношений на них, порожденных, на уровне прозрачности ядер четвертой градации.
Топологическое пространство — множество, в котором определена некоторая система подмножеств — открытых множеств так, что пустое множество и все пространство является открытыми множествами, объединение открытых множеств является открытым множеством и пересечение любых двух открытых множеств является открытым множеством. Множество обобщенных показателей озлокачествления N<i,j>, мощности которых оцифровывают вершины графа, по построению образуют именно такую систему. Можно показать, что при этом топологическое пространство это пара <N<i,j>,I>, где I - операция на множестве N<i,j>, при удовлетворении которой перечисленным условиям, операция I делается "операцией взятия внутренности" (терминология теории множеств).
 Но внутренность каждого множества комплектовалась по построению и определена группировками подмножеств на поверхности частостей распределений ядерно-цитоплазматического отношения. Множество К внутренних подмножеств <N<i,j>> — решетка множеств, на множестве отношений локального гомеостазиса образовало финитную клеточную ситуацию, формально отражаемую аксиоматикой классов состояний.
Но внутренность каждого множества комплектовалась по построению и определена группировками подмножеств на поверхности частостей распределений ядерно-цитоплазматического отношения. Множество К внутренних подмножеств <N<i,j>> — решетка множеств, на множестве отношений локального гомеостазиса образовало финитную клеточную ситуацию, формально отражаемую аксиоматикой классов состояний.
Клеточная ситуация достижения финитного состояния отображается графом формирования внутренности системы локального гомеостазиса, развивающегося в условиях мультиустойчивой стабилизации клеточных потоков на отношениях квантов времени и дискрет информации.
При внимательном рассмотрении легко увидеть, что построение, из вершины пространства характеристической пирамиды центральной перспективной проекции на ее наклонное основание, множеством своих узлов, ветвей и их вершин, образует объемно-пространственную структуру. Все ее элементы либо лежат в разных косых плоскостях, отображающих градации оптической плотности, либо своими связями пересекают их. Инвертируясь относительно направления плоского дерева роста, эта структура приняла положение, при котором отображения множеств клеток отправления и прибытия заняли позиции на различных уровнях градаций прозрачности ядер. На стадиях порождения и развития, множество отправлений клеток баланса эквивалентности занимает нижние позиции, транспортируя клетки в множества прибытия, занимающие позиции на верхних уровнях — уменьшения прозрачности ядер. В соответствии с центральной гипотезой карциногенеза ("Порядок в мире клеточных популяций"), это отражает агрессию возбуждением. На стадии озлокачествления, подключая новые направления клеточного транспорта, множества клеток отправлений формируют подмножества клеток, отражающиеся в баланс толерантности, занимают верхние позиции, а множества прибытия, зарождаясь как подмножества эквивалентности, появляются в позициях нижних уровней. В соответствии с гипотезой, это отражает агрессию затягиванием и отображает появление клеток, втягиваемых в транспортные клеточные потоки.
Множества отправления такого динамического отображения формируются из подмножеств клеток текущих состояний, непрерывно транспортируемых на отношениях дуг графа к последующим узлам — также перетекающим множествам прибытия. Каждое из перетекающих множеств структурировано на множестве своих межклеточных связей и находится в отношениях с другими множествами на сетях групповых межклеточных взаимодействий.
Принципиальная особенность структуры "внутренности" - сцепление множеств клеток различных состояний на 16 вершинах графа, расположенных на различных уровнях пространства топологической пирамиды. Это соответствует уровням клеточных взаимодействий различных подмножеств клеток в их размножающемся множестве и отражает эти взаимодействия. Дуги графа отражают однонаправленность и эквифинальность этих отношений в пространстве субстрата. Дерево финитного состояния отобразилось в объемную решетку множеств. Структура "внутренности" этого множества отражает функции их групповых взаимодействий и направленность клеточного транспорта. Это образует отражение динамических процессов гомеореза — самоуправляемой мультиустойчивой системы управления потоками, как локального гомеостазиса, функционирующего в мультиустойчивой среде ракового гомеостата.
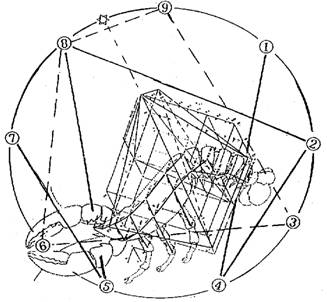
Граф агрессивности локального гомеостазиса определен на плоскости основания характеристической пирамиды центральной перспективной проекцией, вычленяющей на структуре внутренности графа агрессии порождения, развития и озлокачествления топографический контур поверхности, покрывающей пространство сетей коммуникаций (рис.1), очерченный с учетом максимальной погрешности исходных данных +10%. Титульный рисунок (вот он, сюрприз, обещанный Ф.Гродинзом!), отражает вычлененный контур, который воспроизвел естественный образ вида беспозвоночного членистоногого, имя которого было присвоено злокачественной опухоли врачами средневековья. Совмещение контура с девятиугольником граф-октавы метаморфозы биологического субстрата на этом рисунке, позволяет проследить гипотетическое отношение соответствия топографического контура рака как единства структуры и функции, отражающего существование энергетического источника самодостаточности перехода от живого к неживому на клеточном уровне.
Парадоксально, но факт: каждая проекция, из множества проекций вершин графа (структура функциональных подмножеств клеток) и его ветвей (отношений между ними), отразивших решетку множества, почти в точности совпала с тем или иным характерным сегментом или элементом структуры его хитинового корпуса. Так, точка почкования клеток-агрессоров на дереве роста — множества клеток, отраженных балансом эквивалентности, нашла свое отражение в энергетическом, мышечном хвостовом узле существа. Точка роста, отражающая порождение состояния Progression, воспроизвела двунаправленный контур озлокачествления — из пространства третьего сегмента его хвостовой части, опирающийся своими тремя вершинами на пространство трех верхних сегментов этой же части туловища. Контур озлокачествления, в свою очередь отразил порождение, на сегменте изображения туловища членистоногого, левосторонней ветви дерева роста агрессивных множеств клеток-агрессоров. В виде цепочки узлов, это породило дочерние узлы структуры, в свою очередь, отражающие развитие порожденного разрастания множеств клеток, отображенных в членистые конечности двигательного аппарата животного. Фокус целевой функции, как финитная область прибытия, проектируется двумя узлами структуры. Финитные узлы структуры оказываются вне пространства определения оператора, и это показывает, что оператор взятия внутренности образует "двойку" множества и "оператора над ним", не на всем множестве сообщества озлокачествленных клеток опухоли. Из пространства его определения исключается множество прибытия "готового продукта" - подмножества, ставших в процессе канонизации "статическими" голых ядер и пустое подмножество клеток, обладающих превышением площади ядра над площадью всей клетки близким "справа", к значению 0.9: разность мощностей множеств голых ядер и клеток с ядерно-цитоплазматическим отношением 0.9 составляет всего 5%. Это позволяет считать, что полученное отражение финитных множеств есть отображение голых ядер, как готового малигнизированного продукта. Завершив внутриклеточные преобразования в сообществе размножающихся клеток и образовав сообщество раковых жемчужин, они либо выходят из сети информационных межклеточных и групповых взаимодействий динамики клеточного транспорта либо, выполняя до конца целевую функцию локального гомеостазиса, меняют характер своего динамизма и, реализуя новые назначения областей прибытия, отрываются от сообщества и попадают в кровоток или в ток лимфы, но уже в ином функциональном качестве.
Таким образом, утверждение о потере клетками дифференцировки в сообществе опухоли, хотя и фальсифицируется относительно нее, но может быть переадресовано относительно небольшому подмножеству элементов канонического состояния, которые перестали существовать в качестве клеток и действительно приобрели полную автономность в кровотоке или токе лимфы. Пара подмножеств "готового продукта", отразившаяся в оконечные "висячие" множества графа, будучи спроецирована на фигуру злополучного существа, также получает интерпретацию, соответствующую конфигурации контура его каркаса.
Отображение проекции контура поверхности, покрывающей пространство сетей коммуникаций графа агрессии, удивительно не только своей конфигурацией. Численные описатели элементов его архитектуры находятся в отношении соответствия с аналогичными описателями композиции отображенного существа. Именно это усугубляет сложности логической интерпретации предмета парадокса, утверждая объективность и подобность полученного топографического контура конфигурации существа, называемого "раком". Мы столкнулись именно с тем случаем, о котором говорил Ф.Гродинз, имея в виду непредсказуемость появления парадоксов и сложность их интерпретации в количественных терминах.
Совпадения и особенности.
Число элементов хитинового каркаса, очерченных контуром метаморфозы как части тела животного, совпало с их реально существующим числом. Гомеостазическое переключение отображается вершиной, отраженной в "энергетический" узел организма животного, управляющего его хвостовым оперением и это можно трактовать (естественно весьма условно и в рамках парадокса), как отображение состояния самодостаточности. Каждая из вершин графа располагается на изображении одного из характерных элементов этого существа, например — на том или ином сочленении хитинового каркаса его тела. Подмножествам вершин — стратам и фазам состояний, соответствуют отдельные его части. Направления роста в кинетике отображаются на графе по направлению транспорта клеток (в интерпретации фигуры — от "хвоста к голове") по мере возникновения и развития новых коммуникативных сетей, реализующих те или иные межклеточные взаимодействия. Мы, однако, не исключаем, что это совпадение коррелируется с принятым уровнем конкретизации данных исходного описания.
Важным является еще одно обстоятельство. Сопоставление, на титульном рисунке, центральной перспективной проекции топографического контура поверхности, покрывающей пространство сетей коммуникаций графа агрессии с девятиугольником граф-октавы метаморфозы биологического субстрата, позволяет проследить отношения их дуального соответствия. Каждая страта и фаза состояний субстрата, последовательно отображаясь на графе и отражая "свой" элемент конструкции хитинового корпуса существа, находится в отношении динамического соответствия с количеством и порядком одноименных переходов, описывающих отношения порождения на вершинах девятиугольника графа-октавы и соответствует номеру вершины, упорядочивающему ее положение на графе. Вершине 1 соответствует один узел инициации, вершине 2 — два последующих уровня инициации, вершине 3 — три состояния субстрата (INITIATION, PROMOTION, PROGRESSION), вершине 4 — четыре последующие ветви инициации, вершине 5 — пять ветвей цепного роста, вершине 6 — шесть этапов образования голых ядер, вершине 7 — семь ветвей цепного роста (с учетом "ядерного взрыва"), вершине 8 — восемь ветвлений роста, вершине 9 — девять стадий процесса.
Это означает, что структуре полученного контура, подобного телу членистоногого, давшего обобщенное имя "рак" озлокачествленному состоянию и определенной по объективным данным клиники как отображение реальной метаморфозы субстрата, свойственна генеральная упорядоченность 142857 обобщенного отношения "пространство-время". Напомним, что пентаэдр — характеристическая пирамида, в пространстве которой произведены построения, вписывается в единичный шар, в пространстве которого отражаются пространственные фазовые решетки клеточных множеств. Девятиугольник графа-октавы также отражается в этом пространстве Именно это и сделало возможным его использование в качестве инструмента исследования собственных свойств локального гомеостазиса, как ракового гомеостата. Дерево самодостаточного финитного состояния, как отображение единства клеточных, временных и информационных потоков, базируется на тройном отношении соответствия, определенном тремя независимыми путями.
Нормальное и инициальное состояния субстрата, отображенные в пространстве топологии полусферой гомеостазиса, расположенной вне пространств озлокачествления или транспорта, на фигуре членистоногого нашли свое место на уровне хвостовых пластин, также расположенном ниже точки почкования — его энергетического узла и также вне многогранника области определения оператора, в ближайшей наружной окрестности его нижней грани. Область определения оператора I, образующего "двойку" <N<i,j>,I> формального описания топологического пространства, отразилась как область многогранника определения взаимодействий потоков транспорта озлокачествляемых клеток. На полученном графе четко просматриваются отношения общего упорядочения в кинетике процесса. Каждая текущая стадия находит также отображение в "своем" элементе конструкции хитинового корпуса существа, носящего название "рак".
Имеется еще одна его сторона, показавшаяся вначале загадочной, но получившая свою логическую трактовку. Внутренность динамического множества трансформирующихся клеток, формально отразившись в контур, подобный очертанию рака, тут же зародила сомнение: образ членистоногого сформировался без характерных для этого существа клешней — средств его защиты и нападения! Если такое совпадение есть закономерность логического вывода, то это наступательное оружие рака и не могло отразиться в нашей модели. Оно, порожденное творческим воображением, расположилось на рисунке за пределами выпуклого многогранника — области определения рассматриваемого топологического пространства. Логично предположить, что такое отражение "ударного оружия" опухоли как агрессивного новообразования, должно было бы сформироваться как отображение активного характера метастаз на конечном этапе озлокачествления, током крови и лимфы распространяющихся по всему организму, и конечно, отразиться вне этой области, как постфинитный результат процесса трансформации клеточного субстрата.
Но множество клеток субстрата, образующего банк исходных данных, было образовано только по данным обследования поверхностного эпителия, подлежащих слоев (стромы или базального слоя) мышечной и соединительной ткани и каких либо параметров метастаз, полученных из тока крови или лимфы изначально не содержало. Элементы, образующие в биологическом субстрате метастазы, как вторичные патологические очаги, возникающие вследствие переноса их током крови или лимфы из первичного очага озлокачествления, не рассматривались и данные по ним в расчеты не закладывались.
Множество голых ядер сформировало на структуре модели лишь конечную вершину вне пространства определения оператора взятия внутренности, как возможную область отправления метастаз в их "свободный полет". И хотя область их формирования отразилась на контуре той части тела существа, вблизи которой можно себе представить рост клешней, вопрос об отображении метастаз средствами клеточного транспорта становится беспредметным. Но ведь известны и бесклешневые виды этих членистоногих существ!
Парадокс карциногенеза как антиномичное суждение.
Итак, мы можем сформулировать два суждения, находящиеся в отношении противоположности, или даже исключающие друг друга: "финитное состояние локального гомеостазиса, как его модели, отражается в образ членистоногого — "рак", и "весьма сомнительно, скорее всего, что финитное состояние локального гомеостазиса, как его модели, не отражается в образ членистоногого — "рак". Эти суждения производят впечатление, что оба они могут существовать, оба могут быть доказаны в качестве правильных и могут быть конкатенированы в одно высказывание: "финитное состояние локального гомеостазиса отражается в образ членистоногого "рак" и не отражается им." Такое высказывание — антиномия, не обладает строго определенной логической значимостью и, разумеется, оно не истинно. Хотя, с другой стороны, и не каждая из опухолей, имеет образ этого животного!
Кант называл антиномиями те противоречия, которые возникают в случае, когда разум пытается выйти за пределы чувственного опыта и познать "вещи в себе". Мы оказались именно в таком положении. Различают антиномии — логическое отражение противоречий самой действительности и антиномичные суждения — парадоксы, обусловленные конкретным уровнем развития знания. Отражение локального гомеостазиса фигурой рака — первая из них. Отражение незнания, непонимание генезиса полученного совпадения как, собственно, и истинного происхождения самого названия — "рак", как состояния локального гомеостазиса, образует второе.
Обнаружение парадоксов — один из главных источников развития познания. Теория относительности возникла в результате обнаружения антиномичности некоторых исходных положений классической физики. "Противоречие — антиномия", один из видов категории "противоречие" и многие парадоксы образуют ее разновидности. По своему виду это приведенная выше конъюнкция предложения и его отрицания в двузначном исчислении высказываний, но обладающая нечеткой "размытой", fuzzy-логической значимостью. Если ее характеризовать как относительно истинную, то степень относительности может варьироваться в широких пределах и не становиться равной нулю. В случае, если содержательно-конкретное разрешение не удается найти, возникает задача разработки средств, позволяющих временно блокировать парадокс.
В рамках развиваемой теории доказательного объяснения антиномичному суждению о соответствии внешнего образа опухоли как новообразования и образа феномена на морфологии клеточного поля, полученного на основе формальных отношений общей теории систем, не найдено. В поисках средств блокирования с учетом феноменальной сущности предмета парадокса, мы оказались вынужденными искать такое объяснение во внешних относительно развиваемой теории обстоятельствах. В частности, рассматривалась возможность реализации целенаправленного гипотетического внешнего вмешательства в коррекцию эволюции локального гомеостазиса в целях придания ему свойств ракового гомеостата, как средства реализации внутривидовой компоненты "естественного" отбора. Этому рассмотрению способствовали работы доктора технических наук П.Горяева и кандидата технических наук Г.Тертышного в области волновой генетики, показавшие что генетическая информация может существовать в виде электромагнитных полей.
Это единственная из гипотез, которая оказалась не фальсифицированной методами системного анализа. Она, однако, приводит к предположениям, еще более фантастичным, чем сам парадокс: соответствие отраженного в название внешнего образа физической опухоли ее феномену на уровне топологии клеточных множеств, может быть образовано только субъектом внешней среды с высоко развитым разумом, существенно превышающим интеллект фигуранта операции — прародителя человека и обладающим возможностью, с целью задания целевой функции локального гомеостазиса, реализовать коммуникативную функцию на генном уровне. Численные характеристики такого гипотетического вмешательства, в точности совпали с аналогичными характеристиками процесса коррекции эволюции, а инструментарий вмешательства интерпретируется на механизмах волновой генетики, осуществляющих перенос генетической информации в виде электромагнитных полей. Только на генном уровне возможно конструирование структуры рекурсивных управлений размножением клеточных множеств, генерации их математического и программного описания на нестационарных сетях распределенной биологической компьютерной системы. Такой, хотя и кажущийся фантастическим вывод, подтвержденный, тем не менее рядом предварительных исследований, представляя самостоятельный научный интерес, существенно выводит нас за рамки настоящей работы.
И хотя, попав в мир недоумений, мы не имеем уверенных ответов на порожденные вопросы, мы можем сформулировать сентенцию — как мнение о предмете мысли, отражении предмета парадокса в нашем сознании, утверждающее истину. Это мнение может рассматриваться как ответ на вопрос о связи мистики и реальности парадокса:
- суждение, утверждение, интуитивная догадка, даже кажущееся мистическим или мифическим высказывание, как предмет мысли, становится отображением скрытного объекта реальной действительности если, и только если, на множестве отношений этого предмета мысли, может быть установлено число как мера определенности, подтвержденная на видимых проявлениях реального объекта.
Мистика остается мистикой до тех пор, пока остается незнание. А мы, для предмета мысли как предмета парадокса, такое число установили. Но откуда, и каким образом средневековые врачи определили, что название "рак", присвоенное ими злокачественной опухоли, совпадает с ее формальным, на мере определенности отображением, и чем они руководствовались, устанавливая его, мы так до конца и не поняли.
Знание порождает скорбь, незнание умножает ее.
Яков Гельфандбейн
(c) 1011494 CIPO
Популярное



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.