Бигомеостазическое пространство клеточных множеств
 Энергетическое обеспечение процесса озлокачествления
Энергетическое обеспечение процесса озлокачествления
Фазовые сагиттальные оболочки клеточных множеств, образуя замкнутые линейчатые поверхности, отражают границы парциальных направлений роста опухоли в фазовом пространстве, ограничивающие объемы разрастания множеств клеток для этих направлений. Их поверхности отражают ограничения следа траекторий перемещения точек, изображающих изменения выходной координаты процесса целенаправленного управления устойчивым неравновесным ростом в системе локального гомеостазиса — раке.
Оказалось, что фазовая сагиттальная оболочка усредненного, интегрального роста, положением разделяющего слоя, делит пространство состояний на три функциональные подпространства — гомеостазиса, накопления и локального гомеостазиса, в золотом отношении. Существование тройного золотого отношения на множествах трансформирующихся клеток, характеризует естественно-технологический процесс трансформации субстрата как самоорганизующийся и оптимальный.
Плоскость переключающего множества клеток, в этом пространстве, отражает формирование трассирующей сети активации функциональных коммуникаций выходов и входов клеточных компьютерных множеств.
Фазовые сагиттальные подпространства, как составляющие бигомеостазического пространства, образуют множество вторичных отображений, которые позволяют установить функциональные соотношения между их различными представлениями и дать численные оценки процессам клеточного роста на них. В частности, оценки энергетических затрат процесса трансформации показали, что сагиттальные направления роста носят состязательный характер. Отношение состязательности отображает ролевую функцию форсировки процесса инициальным сагиттальным представлением.
Установлено, что запуск механизма целевого управления на клеточной сети изменения состояний, производится не просто при достижении множеством клеток пластического состояния определенной стартовой мощности баланса эквивалентности, но при достижении на этом множестве максимальной скорости активации. Это подтверждает предположение не только о форсирующих свойствах клеток состояния PROMOTION, реализующих вовлечение клеток в процесс трансформации за счет энергетической подпитки извне (феномен ангиогенеза), но и одновременную, подпитку этого процесса за счет собственной, накопленной энергии. Это означает, что множества клеток состояний PROMOTION и PROGRESSION находятся не только в информационно-управляющих, но и в энергетических отношениях. Для уточнения этих, и иных отношений, оказалось необходимым доопределить области отправления множеств клеток исходного описания, соответствующих каждому из выбранных представлений направления роста. Отображение результирующего древесного роста на сети изменения состояния в сеть выхода, отображенного на множестве доопределенных фазовых сагиттальных подпространств, может быть произведено на пересечении соответствующих сагиттальных решеток в пространстве единичного шара и приводит к отражению фазового пространства в пространство топологии.
Фазовая сагиттальная оболочка и ее сужение — пространственная сагиттальная решетка, покрывают множества, определенные на вероятностной мере. Следовательно, изменения объемов оболочек и пространственных решеток по мере роста опухоли отражают, в пространстве времени, изменения их информационного, в шенноновском смысле, наполнения. Процессы роста опухоли, отображаясь в фазовом пространстве, имеют информационный характер.
Все отображения на оболочках сохраняют свойства относительной меры. Поэтому могут быть получены вторичные отображения — мажорантные характеристики оболочек, в виде зависимостей различных подмножеств элементов s множества S текущих состояний, отражающие динамику их порождения. В качестве таких характеристик удобно использовать площадные и объемные отображения различного вида сечений фазовых пространств, в частности — площадей сечений и объемов слоев для различных направлений роста в функции номера слоя на древесном росте.
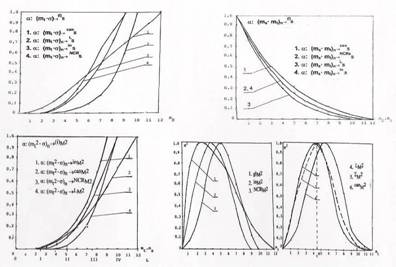 Рисунки отражают функциональную роль целевого управления, как устремление клеточного транспорта в фокус фазового пространства. Отображение произведения редукции времени на рост для каждого слоя (рис.1) отображает поглощение редукции текущего времени относительно момента канонизации, а произведение той же редукции времени на изменение состояния, вырезает временные слои в функции номера фазового слоя и определяет израсходование общего "резерва'' не озлокачествленных клеток относительно их количества, отраженного древесным ростом.
Рисунки отражают функциональную роль целевого управления, как устремление клеточного транспорта в фокус фазового пространства. Отображение произведения редукции времени на рост для каждого слоя (рис.1) отображает поглощение редукции текущего времени относительно момента канонизации, а произведение той же редукции времени на изменение состояния, вырезает временные слои в функции номера фазового слоя и определяет израсходование общего "резерва'' не озлокачествленных клеток относительно их количества, отраженного древесным ростом.
Отображение площадей сечений фазового пространства — произведения редуцированного времени на изменение состояния (рис.2), характеризует остаток незаполненного объема, покрываемого сагиттальной оболочкой. По мере роста, оно отражает изменение мощности множества клеток еще не вовлеченных в рост, и вырезает площади квадратов фазовых слоев в функции нередуцированного роста (на что указывает индекс <n> — изменения роста на рисунке). Остаток незаполненного объема при устремлении роста к фокусу, стремится к нулю.
Отображение произведения квадрата редуцированного времени на рост (рис.3) соответствует заполнению объема фазового пространства, ограниченного соответствующей оболочкой и соответствует росту мощности множества вовлеченных в процесс озлокачествления клеток. С другой стороны, это отражает и характеризует эффективность целевого управления. Некоторые из вторичных отображений связаны отношением обратного соответствия.
На рис.4 приведена еще одна вторичная характеристика. Это — скоростная характеристика процесса озлокачествления, показатель, отражающий скорость нарастания нормированной мощности множества канонизируемых клеток, покрываемых изменяющимся в функции роста объемом того или иного направления роста.
Особенность этого отображения — отражение максимума скорости древесного роста его парциальными составляющими. Разброс времени достижения максимальной скорости роста соответствует более чем трем фазовым слоям — от второго до шестого, а среднее значение соответствует 4.5 фазовому слою. Наиболее "быстродействующий" процесс покрывается оболочкой глобального сагиттального направления роста.
Различия в скоростях и времени процессов клеточных преобразований для различных сагиттальных представлений роста, связаны с долей участия в процессе энергетической подпитки множеств агрессивных клеток состояния PROMOTION, по разному вырезаемых тем или иным сагиттальным представлением. На вторичных отображениях наглядно проявляются и некоторые временные соотношения на процессах, покрываемых фазовыми сагиттальными оболочками.
На титульном рисунке приведена интегральная фазовая сагиттальная оболочка. Эта оболочка отражает покрытие множества таких подмножеств клеток, которые вовлекаются в процесс роста при одновременной и параллельной активации по всем рассмотренным сагиттальным представлениям. Она представляет собой усредненную сагиттальную, состоящую из двух частей, поверхность бигомеостазического пространства, покрывающую суммарные объемы разрастания. Плоскость раздела определяется положением максимума ее скоростной (вторичной) функции (рис.4) и соответствует фазовому слою 4.5: N4÷N5, I3, P3÷P4, end I÷PG0. Стрелка на рисунке указывает плоскость гомеостазического переключения, как верхнюю границу слоя накопления.
Такое положение плоскости раздела может означать, что запуск механизма целевого управления на глобальной клеточной сети изменений, производится не просто при достижении множеством клеток пластического состояния определенной стартовой мощности, но при достижении на этом множестве максимальной скорости активации, что, скорее всего, является следствием форсировок. Характер аналогичных зависимостей множества сагиттальных представлений показывает, что процесс гомеостазического переключения множеств клеток, активируемых агрессивными клетками баланса эквивалентности, "разгоняется" путем последовательного, по всем сагиттальным представлениям (кроме воспринимающего разгон канонического представления роста), с целью сообщения процессу своеобразной "инерции", достаточной для выхода в состояние PROGRESSION с накопленной энергией, и завершения процесса дегенерации клеток.. Это позволяет предположить, что управление гомеостазическим переключением производится по скорости древесного роста. Известно, что такое управление применяется в случаях, когда необходимо использовать накопленную энергию выходной координаты системы. Не исключено, что накопление энергии "инерции", отраженное этими отношениями, связано с обеспечением процесса образования метастаз и их последующим отправлением в токи крови и лимфы.
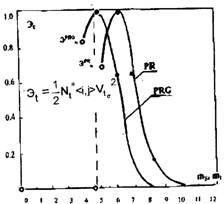 Это предположение не опровергается и с позиций оценки энергетических затрат на возбуждение, поддержание и разгон роста. Можно провести некоторую аналогию с классической механикой (а управление — ветвь этой науки), и ввести в рассмотрение биологический аналог понятия кинетической энергии. Определим кинетическую энергию роста Э на количественной мере трансформации клеточного поля, как половину произведения мощности N множества озлокачествляемых клеток, помноженной на квадрат скорости V размножения клеток этого множества. Так как мощность множества озлокачествляемых клеток была определена в качестве обобщенного показателя озлокачествления, находящегося в отношении соответствия с их физической массой N <i,j>, то энергия роста клеточных множеств запишется в виде соотношения, аналогичному по виду уравнению кинетической энергии в механике. В этом соотношении фигурирует квадрат скорости роста клеточного множества и аналог массы клеточного множества. Такая условная энергетическая характеристика может быть найдена не только для момента переключения, но и для любого момента процесса трансформации.
Это предположение не опровергается и с позиций оценки энергетических затрат на возбуждение, поддержание и разгон роста. Можно провести некоторую аналогию с классической механикой (а управление — ветвь этой науки), и ввести в рассмотрение биологический аналог понятия кинетической энергии. Определим кинетическую энергию роста Э на количественной мере трансформации клеточного поля, как половину произведения мощности N множества озлокачествляемых клеток, помноженной на квадрат скорости V размножения клеток этого множества. Так как мощность множества озлокачествляемых клеток была определена в качестве обобщенного показателя озлокачествления, находящегося в отношении соответствия с их физической массой N <i,j>, то энергия роста клеточных множеств запишется в виде соотношения, аналогичному по виду уравнению кинетической энергии в механике. В этом соотношении фигурирует квадрат скорости роста клеточного множества и аналог массы клеточного множества. Такая условная энергетическая характеристика может быть найдена не только для момента переключения, но и для любого момента процесса трансформации.
На рисунке приведены кривые, иллюстрирующие этот энергетический показатель для ветвей древесного роста PROMOTION, и активируемого (и подпитываемого) множествами клеток этого состояния, основного ствола PROGRESSION. Максимальное значение энергии переключения соответствует изменению состояния и текущему времени 4,5 уровневого слоя. Максимальное текущее значение условной энергии, соответствующей состоянию PROGRESSION, лежит в окрестности этого слоя.
Кривые для обеих состояний достаточно близки, но расход энергии множеством клеток состояния PROMOTION несколько опережает расход энергии трансформирующимися клетками состояния PROGRESSION. Это опережение составляет примерно два межуровневых слоя и подтверждает сделанное ранее предположение о форсирующих свойствах клеток состояния PROMOTION, реализующих вовлечение клеток в процесс трансформации и его подпитку за счет собственной энергии. Это же означает, что множества клеток этих состояний находятся не только в информационно-управляющих, но и в энергетических отношениях.
Для момента гомеостазического переключения вычисленное нормированное значение условной энергии размножения максимально и равно единице. Это свидетельствует о том, что основные (собственные) энергетические затраты множества клеток баланса эквивалентности расходуются на запуск процесса, а его разгон и реализация осуществляются за счет внешней подпитки клеточных множеств состояния толерантности. Этот результат представляется весьма важным с позиций оценки феномена ангиогенеза.
Из этих кривых видно также, что механизм гомеостазического переключения, формируясь на этапе накопления множества переключающих клеток баланса эквивалентности на межуровневых слоях ins1: N2, I1, P1; inS2: N3, I2, P2 и inS3: N4, I3, P3 (идентифицируемых уровнями древесного роста), накопил энергию активации до значения равного 0.65, а энергию, переключения до значения, равного 0.8 от ее максимального нормированного значения. Это энергетически обеспечивает функционирование всех механизмов взаимодействий агрессивности клеток состояния PROMOTION на инициальной сети входов, активизации процесса PROGRESSION на сети входов развитого процесса и поддержания его далее, в устойчиво-неравновесном состоянии вплоть до завершения метаморфозы.
Положение разделяющей плоскости в бигомеостазическом пространстве, ограниченном интегральной фазовой сагиттальной оболочкой, определяется отношением численных отрезков, отражающих текущее состояние субстрата и его редуцированное отражение, а также текущее время и его редуцированное значение, равным 0.618. Это в точности соответствует отношению золотого сечения.
Напомним, что отношение порождения процесса трансформации, отображенное на граф-октаве этим же отношением золотого сечения, отражает двойную оптимальность всего естественно-технологического процесса. В бигомеостазическом пространстве это же отношение порождения, с одной стороны, нашло отражение в положении разделяющей плоскости. С другой стороны, в самой разделяющей плоскости золотого сечения этого пространства, существует еще и "внутреннее" золотое отношение. Оно образуется на подмножествах клеток баланса эквивалентности отношением мощностей подмножеств клеток состояний PROGRESSION и PROMOTION (вычислено с отклонением от него в пределах точности обработки исходных данных). Можно полагать, что такая связь компонент процесса — двойным отношением золотого сечения, проявилась как "закон гармонии систем" синергетики — новой науки о процессах в самоорганизующихся системах. Закон гармонии систем утверждает, что золотое сечение есть числовой инвариант любой самоорганизующейся системы.
Каждый из элементов множества подмножеств на решетке исходного описания входил в свое подмножество вместе с некоторой окрестностью, что было обусловлено методикой сбора подготовки исходного описания. Это означает, что каждый из элементов подмножеств — внутренний. Множество, каждый элемент которого является внутренним — открытое множество. Так как множество подмножеств клеток карциногенеза образует систему открытых подмножеств так, что пустое подмножество и все пространство являются открытыми множествами и их объединение и пересечение также являются открытыми множествами, то множество исходного описания суть топологическое пространство. Понятие топологического пространства будет нами использоваться при изучении системных свойств системы локального гомеостазиса, в частности, в фазовом пространстве.
В заключение необходимо отметить, что для перехода из фазового пространства в топологическое, из объемов, ограниченных сагиттальными оболочками, в силу их свойства мажорантности, должны быть исключены объемы подпространств, в которых аргументы отображения множеств значений обобщенного показателя озлокачествления не определены. Такие объемы определяются характером сагиттального представления, позволяющим в каждом отдельном случае отобразить ограниченные оболочками пространственные сагиттальные решетки, конфигурация которых зависит от представления роста и степени конкретизации исходных данных. В свою очередь это означает, что сагиттальные оболочки должны быть переопределены на решетке характеристического поля таким образом, что область отправления множеств клеток текущего состояния, рассматривается уже не как множество отображений множеств значений показателя озлокачествления для каждого сагиттального представления, но как отображение множества подмножеств клеток исходного описания такое, что для каждого сагиттального подпространства имеет место их отображение в текущее состояние или в рост. При этом пространственная сагиттальная решетка образует сагиттальное подпространство, но определенное уже не на древесном росте, а на топологии исходного описания.
Таким образом, рассмотренное выше понятие сагиттального подпространства (и соответствующей ему оболочки), составляют не более, как методический этап промежуточного описания для перехода к фундаментальному понятию — пространственным решеткам клеточных множеств, отражая лишь некоторую поверхность, ограничивающую пространство отображения роста при том или ином представлении его направления. Именно пересечения таких решеток, в виде примыкающих друг к другу "слоев", форма боковых поверхностей которых определяется характером сагиттальной огибающей, определяют характер функции роста, как управляемой, в системе локального гомеостазиса, выходной координаты на сети выхода множества клеток, вовлеченных в рост, и соответствующую ей целевую функцию. Такие решетки в дальнейшем будем называть пространственными фазовыми решетками.
Отметим еще одно важное обстоятельство: вложение оболочек не просто отображает упорядоченность сагиттальных представлений роста, но определяет также систему межклеточных взаимодействий на сетях групповых коммуникаций между множествами клеток, покрываемыми различными решетками. Такие взаимодействия являются групповыми, по отношению к тем, которые породили то или иное сагиттальное подпространство (решетку) и вторичными по отношению к ним.
Пересечения решеток позволяют оценить функцию роста как след перемещения точки, отображающей изменение выходной координаты в результате именно такого взаимодействия.
Яков Гельфандбейн
(c) 1011494 CIPO
Популярное
Артистка будет одним из членов жюри шоу "Суперстар ВИА".



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.