Фазовые пространства, сагиттальные оболочки и вложения размножающихся множеств
 Целевая функция локального гомеостазиса
Целевая функция локального гомеостазиса
Динамический процесс размножения клеточных множеств как естественно-технологический процесс стабилизации неравновесного роста, в системе локального гомеостазиса может рассматриваться в качестве объекта управления. В терминах теории управления, пространство (множество) в котором определена клеточная управляемая система, определяется как "фазовое" и отображается тройкой множеств — элементов текущих состояний s в пространство состояний S, множества t элементов t текущего времени, определенных на поверхности, "натянутой" на решетку # множества N подмножеств параметров состояния. При этом, каждому из сагиттальных представлений соответствует своя решетка множеств, определяющая вероятный маршрут роста, а следовательно, и свое фазовое подпространство. Фазовое пространство отражает пространственные отношения (взаимное расположение) и формы (геометрические тела) и их обобщения.
Построение фазового пространства удобно производить как суперпозицию парциальных фазовых сагиттальных подпространств. Это позволяет определить их мажоритарные границы в виде набора сагиттальных оболочек — покрытий, отражающих систему множеств, объединение которых, включает множества того или иного представления роста. В каждом таком подпространстве определена траектория — след перемещения точки, отображающей текущее положение выходной координаты процесса, отражающей накопленное изменение состояния биологического субстрата во времени. Траектория этой точки определяет динамику транспортных процессов клеточных множеств для каждого из сагиттальных направлений, а ее усредненное, для множества равновероятных направлений значение, отражает траекторию "генеральной" выходной координаты в предположении маршрутизации процесса одновременно по всем направлениям роста. Как и раньше, роль меры на клеточных множествах фазового пространства, играет предел, влекущий за собой изменение качества объекта в результате накопленных количественных изменений — отношение порождения, выраженное на множестве значений обобщенного показателя озлокачествления.
Знание динамики парциальных составляющих, позволяет изучить транспорт клеток на их генеральном множестве, в котором тем или иным способом определены предельные соотношения, т.е. в топологическом пространстве. Это создает условия для выработки гипотез целенаправленного подавления (срыва) устойчивости процессов неравновесного роста, реализуемых целевой функцией и материализует высказывание W.Ross Ashby о том, что геометрия, обладая собственными силами, "...может точно и последовательно рассматривать разнообразие форм и пространства, далеко превосходящее все, что может дать земное пространство".
С целью минимизации избыточной информации при одновременном повышении наглядности отображения, синтез фазового пространства на множестве сагиттальных направлений роста, был произведен путем конкретизации областей отправления на множестве N подмножеств параметров состояния исходного описания на древесном росте. Области отправления сагиттальных подпространств, формируя фазовое сагиттальное пространство, отражают фактор упорядоченности групповых клеточных взаимодействий транспорта для соответствующих парциальных составляющих.
При этом, каждая из этих областей, отражая одну из возможных составляющих суммарного древесного роста, имеет точкой своего порождения узел сцепления на решетке исходного описания, активируясь коммуникативными взаимодействиями множества подмножеств клеток баланса эквивалентности. Так как порождение этого баланса активирует различные сагиттальные направления роста, то для каждого из них существует и свое сагиттальное представление динамики процесса инициации и подпитки процесса трансформации множествами клеток баланса толерантности и состояния PROMOTION. Такое отображение не обязательно требует рассматривать узел сцепления в качестве активатора, порождающего процесс: оно может производиться, например, из подмножества клеток, принадлежащих эпителию спокойного нормального состояния, отображенного в корень дерева. Такое рассмотрение может оказаться полезным в задаче уточнения фазового пространства раннего озлокачествления на уровнях роста, предваряющих возникновение баланса эквивалентности. Это, однако, требует повышения степени конкретизации, и влечет за собой необходимость уменьшения шага дискретности (увеличение числа отсчетов) ядерно-цитоплазматического отношения и соответственно — увеличения наблюдаемого числа градаций прозрачности ядер клеток.
 Построение фазовых сагиттальных пространств связано с рядом принципиальных трудностей. Одна из них — сложность выбора удобной для одновременного отображения всех компонентов процесса системы координат и их отсчета, другая — невозможность фиксации текущих временных параметров или опорных моментов времени. Последнее порождается ранее упоминавшейся спецификой наблюдения динамики морфологической картины клеточного поля.
Построение фазовых сагиттальных пространств связано с рядом принципиальных трудностей. Одна из них — сложность выбора удобной для одновременного отображения всех компонентов процесса системы координат и их отсчета, другая — невозможность фиксации текущих временных параметров или опорных моментов времени. Последнее порождается ранее упоминавшейся спецификой наблюдения динамики морфологической картины клеточного поля.
Для построения фазового пространства, в связи с нормированием значений параметра озлокачествления, оказалось удобным использовать специфическую систему отсчета, вписанную в объем единичного шара. В этом объеме, пространство определения роста, текущего состояния множества трансформирующихся клеток и времени, ограничивается объемом, вырезаемым поверхностью оболочки, ограничивающей конкретное сагиттальное пространство (прибытия) в объеме прямоугольной призмы единичной высоты с единичной диагональю, опирающейся на диаметральное сечение единичного шара и расположенной в первом квадранте шарового пространства. Во избежание чисто технических трудностей изображения секущих поверхностей и внутренних пространств на иллюстрациях, мы используем его незначительный (против часовой стрелки) разворот, представивший возможность добиться, для того или иного сагиттального представления, наглядности достаточно сложных графических конструкций внутри шара. Такой разворот допускается нормированностью значений параметров состояния, приводящей лишь изменение масштаба проекций на плоскости системы координат.
В результате построений образовались две (геометрически) равновеликие треугольные призмы с общей гранью — вертикальной плоскостью на биссектрисе основания системы координат, рассекающей фазовое пространство на подпространства состояний и текущего времени.
Оцифровка шкалы роста вдоль оси ординат, соответствует трехмерному росту в нормированных единицах сложности уровней древесного роста. Количество уровней в границах нормированного (единичного) интервала, зависит от характера сагиттального представления так, что количество межуровневых интервалов и их длина могут быть различными, отличаясь в зависимости от значения сложности вершин дерева, и от направления роста для различных сагиттальных представлений. Это, справедливо и для изменения межуровневых и межстратовых состояний (переходов).
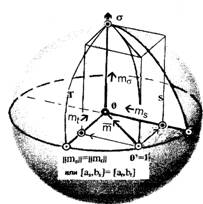
Таким образом, ребра прямоугольной призмы образуют систему координат с осями отсчета роста (ординат), изменения состояний m (абсцисс) и текущего времени t (аппликат), как правила, по которому точкам пространства ставятся в соответствие их координаты с неравновеликими интервалами отсчета.
Отсчет изменения состояния производится по положению модуля вектора m, лежащего в секущей плоскости призмы, который перемещается параллельно плоскости основания системы координат в направлении роста, вдоль оси ординат. Его перемещение образует вертикальную линейчатую поверхность, верхняя грань которой, отражая изменения состояния длиной вектора, образует фазовую траекторию.
Отсчеты по идентификаторам состояния производятся по проекциям фазовой траектории на координатные плоскости, как редукции текущего состояния и времени, а по нормированному значению роста — по меткам на оси ординат. Направление отсчета — от фазовой траектории и ее проекций к оси ординат, как к оси притяжения.
Таким образом, две примыкающие прямые треугольные призмы фазового пространства, ограничивают подпространства отображения роста, изменения состояний и времени. Верхнее основание обеих подпространств образуется некоторой линейчатой, вписанной в объем прямоугольной призмы, криволинейной поверхностью, вырождающейся, в каждом из подпространств в точку, образуя фокус. Эти поверхности, со взаимно перпендикулярными образующими, параллельными основанию координат, и вырезает из объема прямоугольной призмы соответствующее фазовое сагиттальное подпространство. Изменения состояний и редукции времени, помеченные номером уровня, отображаются их идентификаторами на пересечении линейчатых поверхностей, вырезающих объем сагиттального подпространства. Эти пересечения образуют фазовую траекторию изменения состояний, отраженную в формы и объемы соответствующих пространств.
 Порядок следования элементов направленных множеств изменения роста, состояния и текущего времени, изображается в редуцированной, относительно начала координат 0, системе отсчетов, помеченных идентификаторами. В силу направленности этих множеств, предельные рост, состояние и редуцированное время, всегда стремятся в область прибытия, как к устойчивому фокусу. Их начальные значения, как области отправления, могут отсчитываться от спокойного состояния нормального эпителия или с момента гомеостазического переключения, или от любого произвольно выбранного текущего состояния.
Порядок следования элементов направленных множеств изменения роста, состояния и текущего времени, изображается в редуцированной, относительно начала координат 0, системе отсчетов, помеченных идентификаторами. В силу направленности этих множеств, предельные рост, состояние и редуцированное время, всегда стремятся в область прибытия, как к устойчивому фокусу. Их начальные значения, как области отправления, могут отсчитываться от спокойного состояния нормального эпителия или с момента гомеостазического переключения, или от любого произвольно выбранного текущего состояния.
Нумерация меток текущего положения изображающей точки вдоль траектории ее перемещения по времени, соответствует уровням состояний и производится по направлению от периферии к фокусу. Расстояния между метками определяются числовыми криволинейными отрезками такими, что их нормированные величины могут иметь различную длину. Рост, изменения состояния и интервалы времени, отображаются замкнутыми множествами поименованных меток, т.е. множествами, содержащими все свои предельные точки (накопления множеств) и следовательно, фазовые сагиттальные подпространства образуются как замкнутые.
При соблюдении сформулированных условий построения, ось ординат отражает в выбранном пространстве роль оси притяжения, образуя фокусирующую линию, к которой стремятся все элементы множества текущих состояний. Фокус образуется как особая точка фазового пространства. В терминах теории управления общей теории систем — этот фокус — абсолютный, отображающий целевое направление устойчивого движения, в данном случае — устойчивого неравновесного изменения состояний — трехмерного роста. Фокус образует элемент притяжения для всего множества траекторий сагиттальных представлений процесса трансформации.
Устремление состояния множеств клеток биологического субстрата, динамически изменяемого в физическом пространстве и времени в направление фокуса, независимо от характера маршрутизации процесса активации, отражает сущность целевой функции локального гомеостазиса, реализуемой на сети изменений биологической компьютерной системы.
Единичная сфера для фазовых сагиттальных подпространств, находясь в отношении соответствия с осью притяжения, играет двоякую роль. На участке гомеостазиса и накопления переключающего множества клеток, начиная с узла инициации, она оттягивая фазовую траекторию от поверхности основания системы координат, одновременно отталкивает ее от себя и направляет к оси притяжения. В момент гомеостазического переключения она, изменяя характер, приобретает свойства сепаратрисной поверхности, и играет роль элемента притяжения фазовой траектории к фокусу. Однако, даже для суммарного древесного роста, фазовая траектория не совпадает со сферой, стартуя от нее своей изображающей точкой узла инициации и имея с ней касание лишь в фокусе, что образует предельные отношения. Это придает единичной сфере характер ролевой функцию критерия, мерила качества целевого управления: ее поверхность образует систему отсчета качества сохранения локального гомеостазиса с критериальной поверхностью, равноудаленной от начала системы координат фазового пространства, совпадающего с собственным центром. Последнее определяет независимость качества контроля от состояния субстрата и времени
Исходя из высказанных предпосылок и договоренностей, было построено редуцированное фазовое подпространство для каждого из сагиттальных представлений направления роста. След траектории текущего положения изображающей точки, в соответствии с договоренностью, отображается в нем на секущей плоскости фазового пространства началом вектора изменений, лежащим на траектории, а конец направлен в точку текущего роста на оси притяжения. Вектор изменений перемещается вдоль оси притяжения в направлении абсолютного фокуса, параллельно биссектрисе координатного угла системы координат. Этот же след может отображаться и в системе полярных координат, положением конца радиуса — вектора, вращающегося против часовой стрелки от начального положения, совпадающего с основанием системы координат, до абсолютного фокуса, также лежащего на поверхности единичной сферы.
Таким образом, фазовое пространство локального гомеостазиса для любого из сагиттальных представлений, как и процесса в целом, отображается в замкнутом единичном шаре, но оболочка любого фазового сагиттального подпространства, будучи вписана в единичную сферу, сама не является ею, и образует мажорантную поверхность. Ни один из векторов состояний из их множества, порождающего след траектории изображающей точки, кроме начального и конечного векторов, не касается ее поверхности. Это — предельные векторы исходного состояния и конечного изменения.
Уровни состояний образуют в фазовом пространстве уровневые слои, вырезаемые из единичного шарового слоя, параллельные плоскости основания системы координат, и определенные на квадратах основания прямоугольной призмы со сторонами, отражающими текущие состояния m и времени t. Плоскости сечений несут разнообразную информацию о текущих состояниях субстрата, а боковая вертикальная поверхность слоя, своей толщиной и формой образующей, отображает послойное изменение состояния — послойный клеточный переток, и характеризует его. Оболочка фазового пространства, образующая поверхность того или иного сагиттального направления роста, именуется его именем.
Понятие фазовой траектории позволяет детализировать концепцию целевой функции локального гомеостазиса, поставив ее в зависимость от характера принятой системы отсчета координат:
- целевая функция локального гомеостазиса отражается как механизм управления, перемещающий в секущей диаметральной плоскости единичного шара вектор состояния так, что в результате суперпозиции всех типов сагиттальных направлений роста, абсолютное значение редукции роста и изменения состояния минимизируется относительно исходного состояния инициализации, определяемого значением радиуса-вектора равным единице, до его конечного изменения, при котором он стремится к нулю, причем множество изменений в интервале от исходного состояния до конечного изменения, образует максимальное совершенное подмножество.
Для фазового пространства, отображенного в полярной системе координат, целевая функция локального гомеостазиса отражается как механизм управления, вращающий против часовой стрелки, в той же секущей диаметральной плоскости единичного шара, радиус-вектор изменения состояний, направленный из его центра 0 на образующую сагиттальной оболочки — фазовую траекторию движения изображающей точки так, что в результате параллельного и совместного осуществления всех типов сагиттальных направлений роста, он повернется на угол в 90 градусов, причем множество изменений на том же интервале, также образует максимальное совершенное подмножество.
Другими словами: целевая функция, как инструмент механизма управления, направляет изображающую текущее состояние трансформирующегося субстрата точку, к фокусу фазового пространства из любых текущих состояний по всем доступным направлениям, сохраняя заданное качество управления относительно критериальной поверхности, равноудаленной от начала отсчета. Есть все основания полагать, что такое управление реализуется как оптимальное. Это позволило дать формальное определение фазового пространства методами общей теории систем.
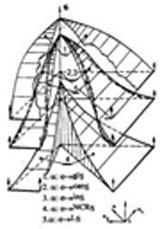
На рисунке, в качестве примера, показаны две практически совпадающие по конфигурации сагиттальные оболочки, определенные для сагиттальных направлений роста, приведенных на том же рисунке. Сопоставление этой, и других пар оболочек, позволяет высказать ряд обобщающих и частных суждений.
Оболочки сагиттальных представлений находятся в отношении сходства, и образуют линейчатые поверхности с горизонтальными образующими, лежащими под прямым углом друг к другу, каждое конкретное положение которых, отражает текущие рост и состояние субстрата, на плоскостях состояния и времени собственной системы координат. Пересечение линейчатых поверхностей на плоскости биссектрисы координатного угла, образует криволинейную фазовую траекторию, соответствующую линии перетока, в свою очередь лежащую на линейчатой поверхности с вертикальной образующей. Форма изменения траектории отражает оптимальность перетока. Перетоки клеток отражают феномен озлокачествления.
Особенностью системы отсчета фазового пространства, является неравновеликость числовых отрезков, образующихся в виде интервалов на осях системы координат, и отражающих нормированные значения роста, изменения состояний и текущего времени. Эта особенность, для различных направлений роста, отражает их относительное отставание или опережение, зависящее от характера переходных процессов форсировок, как реакций активируемых клеток на возбуждения их подмножествами агрессивных клеток состояния PROMOTION баланса эквивалентности. Эта особенность отражается также в различие идентификаторов вдоль фазовой траектории, номеров и состава уровней состояния, при перемещении образующих линейчатых поверхностей, а также в количество числовых отрезков для каждого из направлений роста. Отмеченные отличия иллюстрируются таблицей идентификаторов состояний, приведенной на рисунке.
Форсирующие свойства ряда подмножеств клеток инициального представления относительно канонического, отражаются с опережением в 2-3 уровня роста на инициальном сагиттальном представлении. Это означает, что маршрутизация активации по этому сагиттальному направлению роста, завершает процесс инициации на девятом уровне, переходом в состояние PROMOTION и PG4, форсируя и обеспечивая устойчивую завершенность текущего состояния PROGRESSION. Завершение процесса для канонического направления, отражается в тринадцати уровнях роста дегенерацией клеток и образованием голых ядер. Опережение обуславливается энергетической поддержкой устойчивости роста (неравновесности состояния) на десятом и последующих завершающих слоях канонического сагиттального представления. Скорее всего, это следствие усиленного кровоснабжения агрегированных множеств клеток состояния PROMOTION и PG4 (феномен ангиогенеза).
Процессы трансформации, отображаемые этими (и другими) сагиттальными представлениями, носят состязательный характер, при котором распространение процесса по инициальному сагиттальному представлению играет ролевую функцию форсировки, а множества клеток, активирующих этот процесс, выполняют, по терминологии общей теории систем, роль форсирующего элемента. Срыв форсировок прерыванием энергетической поддержки, может стать эффективным средством торможения процесса.
Принцип "форсировки", результативный в системах искусственных технологий автоматического управления, проявился и оказался решающе важным на великом феномене жизни. Опережение, порожденное влиянием клеток баланса эквивалентности состояния PROMOTION, отображает явление "возбуждения" активации, как фактора проявления агрессивных свойств множества клеток этого состояния и характеризует результативность целевой функции, как агрессивного средства управления.
На оболочке фазового инициального подпространства помечен слой ins4. Этот слой соответствует моменту гомеостазического переключения в этом пространстве, и определяется как конкатенация фазовых слоев N5, P4, PG0, end I. Для канонического сагиттального представления, этому же моменту гомеостазического переключения, соответствует отстающий от него слой s5. Это отражает нарастание форсировки процессом инициализации, что может быть, как и ранее, следствием усиления энергетической подпитки множества агрессивных клеток этого слоя и увеличения мощности множества клеток, вовлекаемых в процесс роста. Гомеостазические переключения на различных направлениях роста, происходят на различных слоях состояний. Это свидетельствует о соревновательности переключений с одной стороны, и о временной протяженности переключающего скачка, с другой. Это также подтверждается значениями параметров пространства накопления переключающего клеточного множества.
Пространство накопления отразилось в результате построения фазовой траектории клеточного транспорта из спокойного состояния нормального эпителия (N0), как исходного, в множество клеток баланса эквивалентности. При этом, исходное подпространство нормального эпителия разделилось на два под(под)пространства: первое из них определяет пространство гомеостазиса, второе — порождения и накопления мощности множества активирующих клеток этого баланса. Порождение гомеостазического перехода отражается верхней поверхностью слоя клеточного баланса эквивалентности ins2 или S3 (N3, i2,P2) (примыкающей поверхностью слоя S4) и верхней поверхностью слоя канонического пространства ins4 или S5 (N5,P4,PG0,endI) (примыкающей поверхностью слоя S6) соответственно. Это — переключающие поверхности маршрутов активации.
Таким образом, в фазовом пространстве отображается три его функциональные подпространства: гомеостазиса, накопления переключающего множества и локального гомеостазиса, разделенные переключающим слоем, верхняя поверхность которого отображает переключающую поверхность. Это образует бигомеостазическое фазовое пространство, охватывающее все возможные, в том числе — переходные состояния биологического субстрата. Именно в этом пространстве отражаются антагонистические феномены гомеостазиса и локального гомеостазиса, а также отношения транспорта между ними. На бигомеостазическом фазовом пространстве, отражаются различные характеристики гомеостазического переключения.
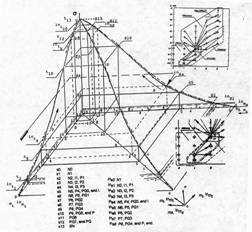
Сагиттальные покрывающие оболочки различных направлений роста, имеют аналогичный характер, собственную форму и собственные характеристики. Элементы множества сагиттальных оболочек находятся в отношении инъективного соответствия — они образуют вложение. Вложение отражает отношение соответствия между множествами клеток различных представлений направления роста, при которых различным элементам из множества состояний одного представления, соответствуют различные элементы из множества состояний других представлений. Рассмотрение вложения (титульный рисунок) показывает, что оболочки парциальных составляющих роста ограничены пространством, имеющим общее основание на прямоугольнике исходного описания с единичной диагональю, сторонами отображающим интервалы и уровни состояния и времени, и общей для всех оболочек точкой — устойчивым фокусом, лежащим на единичной сфере. Вложения образуют систему предпочтений на отношении следования по объемам покрываемого оболочками подпространства из вырезаемого ими фазового пространства, — мощностям покрываемых клеточных множеств. На рисунке, для наглядности, оболочки разделены так, чтобы показать характер вложения и форму фазовой траектории для различных сагиттальных направлений роста. Имея мажорантный характер, сагиттальные оболочки отражают верхнюю границу мощности покрываемых клеточных множеств. Множество объемов образует последовательность, находящуюся в отношении предпочтения.
Особенность системы предпочтений мощностей покрываемых сагиттальными оболочками клеточных множеств, состоит в том, что предпочтениям объемов, соответствуют предпочтения не просто мощностей покрываемых ими множеств клеток, но их подмножеств, группировки которых в множества, классифицированы по парным соответствиям значений аргументов (ядерно-цитоплазматического отношения и прозрачности ядер) обобщенного признака озлокачествления N на решетке исходного описания. Группировки определяются характером сагиттального представления, как отражением вероятного направления роста.
Это означает, что системе предпочтений объемов, покрываемых фазовыми оболочками, соответствует система отношений мощностей N множеств подмножеств клеток, обладающих различными сочетаниями парных значений аргументов обобщенного показателя озлокачествления.
Рассмотрение системы предпочтений позволяет еще раз подтвердить уже высказанное ранее мнение, что развитие процесса трансформации определяется не просто мощностью множества вовлеченных в процесс озлокачествления клеток, но агрессивностью активирующих клеток состояния PROMOTION, образующих множество баланса эквивалентности и форсирующих, в условиях их энергетической подпитки, процессы PROGRESSION. Эти процессы завершаются дегенерацией клеток и покрываются оболочкой канонического состояния, находящейся в отношении эквивалентности с оболочкой инициального сагиттального направления.
Клетки, образующие множество баланса эквивалентности — первоочередные объекты для целеполагания в системе суперселективного клеточного репрессирования.
Как это видно из вложений, отраженных системой предпочтений, минимальное множество вовлеченных в процесс озлокачествления клеток, свойственно сагиттальному направлению роста, отражающему изменения прозрачности ядер. Это может отражать слабую информативность самого признака, как результата слабой реактивности оптических характеристики ядра на биофизические процессы, происходящие в нем при озлокачествлении.
Яков Гельфандбейн
(c) 1011494 CIPO
Популярное
Супруга Евгения Петросяна Татьяна Брухунова сфотографировалась в яркой юбке. Знатоки вычислили стоимость наряда жены юмориста.



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.