Организация топологического пространства локального гомеостазиса
 Сумма технологий активации и агрессивности клеточных множеств
Сумма технологий активации и агрессивности клеточных множеств
Мы различаем математическое пространство (топология) и пространство как порядок и протяженность материальных объектов. При этом, пространство рассматривается как продукт восприятия, из которого вычленяется транспортное подпространство, в котором определена сеть. Эти пространства находятся в отношениях взаимного соответствия. Рассматривая абстрактное фазовое пространство, мы отображаем его в пространство, на котором определен порядок материальных элементов — клеточных множеств.
Множество атрибутов, определяющих множество отношений на множествах элементов топологического пространства локального гомеостазиса, включает подмножества отношений межклеточных взаимодействий активации и агрессивности. Подмножества клеток, находящихся в отношениях активации, агрессивности и порождения процесса озлокачествления, отражаются в топографию гомеостазических шаров. Текущие свойства и параметры шаров отражают динамику их функциональных ролей. Организация топологического пространства и отношений между элементами его архитектуры показывает, что на ней существуют, определенные на мере озлокачествления Гомпертца G, пространства агрессивности. Этим пространствам свойственно четкое разделение подмножеств клеток по выполняемой ролевой функции. Это означает, что система агрессивных клеточных подмножеств локального гомеостазиса находится в управляемых отношениях согласования функциональных взаимодействий агрессивности. Характеристики пространств агрессивности являются либо динамичными, либо инвариантными относительно преобразований, связанных с ростом. Инварианты можно рассматривать в качестве параметров системы локального гомеостазиса как системы, стабилизирующей неравновесный рост.
Основные динамические показатели процесса и множества всех изменений состояний как объекта управления, мы проанализировали ранее и для этого нам пришлось пройти путь от рассмотрения сагиттальных направлений активации до пересечений пространственных фазовых решеток. Мы также выявили существование некоторых разделяющих функций на характеристической плоскости карциногенеза, определенных этими пересечениями. Однако, в структуре отношений, кроме рассмотренных, существуют и иные, вторичные разделяющие составляющие и порождаемые ими вторичные функциональные отношения. Эти отношения не просто сохраняют, но усиливают агрессивность клеточных множеств, реализующих достижение целей стабилизации роста, обуславливая функциональную совместимость и суперпозицию информационной и исполнительной технологий.
Рассмотрим пространства агрессивности локального гомеостазиса. В качестве меры отображения отношений на системе множественных межклеточных взаимодействий, используем понятие меры озлокачествления G.
Геометрические отношения.
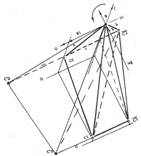
 На рисунке 1 показана проекция прямого сечения единичного шара фазового пространства локального гомеостазиса со своей системой координат, имеющей начало в точке 0 — центре шара. Центры шаров гомеостазиса С1 и локального гомеостазиса С2 лежат на одном и том же уровне озлокачествления, отраженном в значение радиуса шара гомеостазиса R1, но в разных плоскостях — соответственно в плоскости II и III градаций прозрачности ядер. Центры обеих шаров гомеостазиса С1 и локального гомеостазиса С2 и их проекции, в диаметральных плоскостях соответствующих сфер (обозначения приведены на рисунке), соединены между собой и началом 0 системы координат отрезками прямых. Линии, соединяющие центры С1 и С2 шаров и их проекции С'1 и С'2 на собственные верхние полусферы, лежат в первом квадранте единичного шара, в пространстве между плоскостями указанных градаций прозрачности ядер. Линия, соединяющая проекции центров шаров на нижнюю полусферу, проходит в его четвертом квадранте, ее проекция на плоскость основания координат х0L параллельна линии С1С2 и также пересекает пространство между II и III градациями прозрачности ядер.
На рисунке 1 показана проекция прямого сечения единичного шара фазового пространства локального гомеостазиса со своей системой координат, имеющей начало в точке 0 — центре шара. Центры шаров гомеостазиса С1 и локального гомеостазиса С2 лежат на одном и том же уровне озлокачествления, отраженном в значение радиуса шара гомеостазиса R1, но в разных плоскостях — соответственно в плоскости II и III градаций прозрачности ядер. Центры обеих шаров гомеостазиса С1 и локального гомеостазиса С2 и их проекции, в диаметральных плоскостях соответствующих сфер (обозначения приведены на рисунке), соединены между собой и началом 0 системы координат отрезками прямых. Линии, соединяющие центры С1 и С2 шаров и их проекции С'1 и С'2 на собственные верхние полусферы, лежат в первом квадранте единичного шара, в пространстве между плоскостями указанных градаций прозрачности ядер. Линия, соединяющая проекции центров шаров на нижнюю полусферу, проходит в его четвертом квадранте, ее проекция на плоскость основания координат х0L параллельна линии С1С2 и также пересекает пространство между II и III градациями прозрачности ядер.
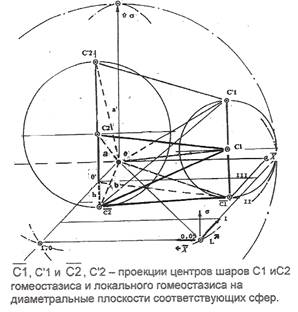
Пространственные отношения полученного построения хорошо видны в иной проекции, на рисунке 2. Отрезок прямой, соединяющий центр шара гомеостазиса С1 и проекцию центра шара локального гомеостазиса на его нижнюю полусферу, лежит в секущей плоскости, между плоскостями II и III градации прозрачности ядер. Удаление проекции центра шара локального гомеостазиса на его нижнюю полусферу от центра шара гомеостазиса С1 отражает множество всех отображений изменений — каноническое состояние субстрата. Наклонная плоскость, проходящая через центр единичного шара 0, в которой лежит прямая, соединяющая центр С1 шара гомеостазиса с проекцией центра С2 шара локального гомеостазиса, рассекает пространство полученного пентаэдра на две части. Эти части образуют две косые трехгранные пирамиды с общей гранью и общей вершиной 0. Будем эти пирамиды и образуемый ими пентаэдр, называть "малыми". Отрезки прямых, соединяющие между собой и с центром 0 единичного шара проекции радиусов гомеостазических шаров на обе полусферы, образует большой пентаэдр с вершиной 0 в центре единичного шара и основанием, образованным как продолжение основания малого пентаэдра. Секущая плоскость большого пентаэдра, аналогично разделяет его на две большие трехгранные пирамиды.
 Плоскости 0IV градаций прозрачности ядер клеток, вырезают в объеме единичного шара шаровые слои. Плоскость IV градации прозрачности ядер лежит в диаметральной плоскости единичного шара на плоскости основания системы координат. Шаровые слои 0 и I градаций прозрачности ядер, оказываются не нагруженными и точек, отображающих множества клеток, участвующих в процессе озлокачествления, не несут.
Плоскости 0IV градаций прозрачности ядер клеток, вырезают в объеме единичного шара шаровые слои. Плоскость IV градации прозрачности ядер лежит в диаметральной плоскости единичного шара на плоскости основания системы координат. Шаровые слои 0 и I градаций прозрачности ядер, оказываются не нагруженными и точек, отображающих множества клеток, участвующих в процессе озлокачествления, не несут.
Рассмотрим отношения на полученных отображениях, и выразим их на мере G озлокачествления.
Характеристическая пирамида агрессии.
Рисунок 2 построен разворотом пентаэдра, показанного на рис.1, вместе со связанной с ним системой координат, против часовой стрелки относительно оси редукции изменения состояний х на 90º, таким образом, чтобы "опереть" радиус шара гомеостазиса на плоскость II градации прозрачности ядер, как на ребро малого пентаэдра. Плоскость в которой лежат центры обеих шаров С1, С2 и их проекции на соответствующие сферы, в соответствии с их взаимным расположением, опирается своими ребрами на плоскости II и III градаций прозрачности ядер и пирамида таким образом оказывается наклонной. Плоскости градаций прозрачности ядер L, как и ранее, образуют шаровые слои с сохраненной нумерацией, начало системы координат и поверхность IV градации прозрачности ядер, остаются на диаметральной плоскости, но развернутой на 90о. Полученный многогранник с вершиной в 0, замыкает пространство межклеточных взаимодействий множества подмножеств покрываемых им клеток, как непустого связного открытого множества точек топологического пространства порождения и развития древесного роста, отраженного в объем единичного (относительно редукции изменения состояний х) куба и образует в его объеме характеристическую пирамиду. Верхняя и нижняя грани куба образуются плоскостями IV и 0 градации прозрачности ядер клеток, вырезающими шаровые слои единичного шара, боковые — диаметральными плоскостями шара, как и ранее играющими роль координатных поверхностей изменения состояний и роста. Древесный рост клеточного множества в характеристическом пространстве пирамиды может быть отображен пространственным трехмерным графом, время наблюдения которого является параметром.
Топография множества подмножеств вершин пространственного графа образует характеристическую решетку единичного куба. Реализация клеточного транспорта в форме распределения подмножеств клеток на узлах этой пространственной решетки, порядок их порождения и динамизмы изменения их мощности, отображают динамику перетоков и транспорта клеточных множеств в пространстве изменения состояний. Изменения состояний, отражаются на этой решетке графом агрессивности, как результатом совокупного, накопленного воздействия факторов агрессивных взаимодействий, порождающих процесс. Изменение фактора агрессивности, выраженное на мере Гомпертца, отражает направление и эффективность активирующих клеточных взаимодействий. Граф агрессивности может быть построен по данным клиники для любого замороженного состояния субстрата
Граф агрессивности локального гомеостазиса.
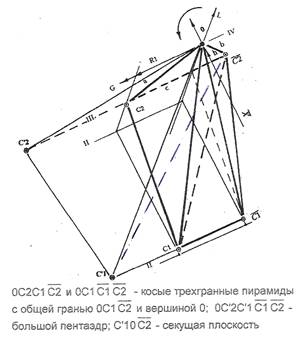
На рисунке 3 показан граф агрессивности активации, порождения и развития, отражающий завершенный процесс достижения биологическим субстратом канонического состояния. Дуги графа, проведенные одной линией, отражает ветви подмножества вершин состояния PROMOTION ACTIVE, двойной — ветви состояния PROGRESSION. Важнейшая особенность графа — его трехмерность в объеме характеристической пирамиды.
Из структуры графа видно, что на пространственной решетке характеристической пирамиды отображается часть общего процесса, не содержащая подмножеств клеток состояния INITIATION, лежащих на плоскостях 0 и I градации прозрачности ядер и подмножеств клеток, образовавшихся после ракового взрыва. Клетки подмножеств, следующих за раковым взрывом, отражаются в плоскости IV градации прозрачности ядер, полностью реализовав агрессивную целевую функцию локального гомеостазиса. Они вышли за пределы его сферы на некотором множестве, вычлененном на архитектуре топологического пространства.
Таким образом, граф агрессивности формируется, начиная с узла сцепления, на тех уровнях страт состояний, на которых развиваются активируемые множеством клеток баланса эквивалентности состояния PROMOTION ACTIVE - с одной стороны, и реализуемые в результате активации процесса, множества клеток страт состояния PROGRESSION, PROGRESSION VITAL - с другой. При построении графа, в качестве показателя, численно отражающего фактор агрессивных взаимодействий, использовалась мера Гомпертца G, определенная для соответствующих уровней озлокачествления на отображении древесного роста. Отсчет этой величины (рис. 2) производился от плоскости "редукция состояния х — прозрачность L ядер" системы координат, полученной после разворота, причем приросты суммировались по правилу векторов. Начальная вершина процесса озлокачествления, соответствующая на характеристической плоскости узлу сцепления <0.3,II>, расположена на плоскости II градации оптической плотности ядер клеток, в сечении, соответствующем значению NCD=0.3 ядерно-цитоплазматической константы.
Полученный граф отражает изменение состояния субстрата, как результат функционирования системы совокупных, совместных и согласованных межклеточных информационных и исполнительных технологических взаимодействий, результат которых отображается в парные значения редукции состояния х и прозрачности L ядер на значениях показателя озлокачествления N<x,L>.
Сумма технологий, отраженная на графе, позволяет высказать ряд важных соображений.
- а. Плоскость, секущая внутреннее пространство пентаэдра, играет роль внутренней характеристической разделяющей поверхности. Объем, заключенный в большуюправую трехгранную пирамиду покрывает множество вершин графа * {<0.4x>0.2;II<L<III} состояния PROGRESSION на плоскостях II и III градаций оптической плотности, появившихся в результате агрессии, а также вершин + множества {<0.3x0.4>;LIII}, активировавших их появление из состояния PROMOTION ACTIVE. Стрелки при дугах графа указывают направление активации, а его структура уподобилась пространственному "куполу", напоминающему плакучую иву, и охватывающему вершины, лежащие на нижних слоях, содержащие множества клеток, подвергшихся активации
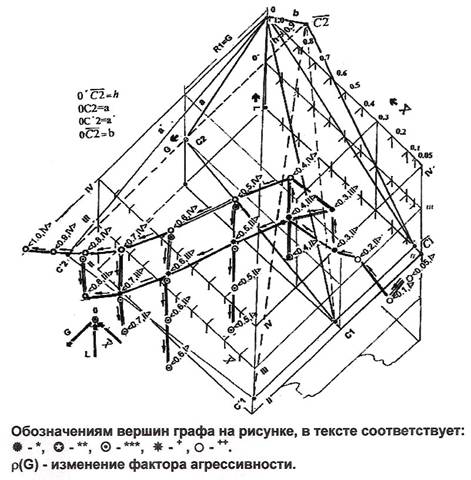 - b. Объем, заключенный в большую левую трехгранную пирамиду покрывает множества {<0.5x0.8;IIILIV>} вершин ***, принадлежащих состоянию субстрата PROGRESSION, появившихся в результате разрастания клеточных множеств, как эффекта активирующих воздействий и совпадающих с ними вершин + состояния PROGRESSION на плоскости III градаций прозрачности ядер. Множество {<0.5x1.0;L=IV>} вершин ** дерева подсостояния PROGRESSION VITAL, пересекло наружную грань C1С20C1 малого пентаэдра, и аналогично множеству клеток ** {<0.4=x>;L=IV} состояния PROMOTION ACTIVE, занимает положение вне малого характеристического пентаэдра на плоскости четвертой градации прозрачности ядер, внутри большого пентаэдра. Множество вершин *** {<0.5x0.8,ILII>}, активируемых агрессивными подмножествами клеток * состояния PROGRESSION, также находится вне объема пентаэдра, в объеме единичного куба на плоскости II или даже I градации прозрачности ядер.
- b. Объем, заключенный в большую левую трехгранную пирамиду покрывает множества {<0.5x0.8;IIILIV>} вершин ***, принадлежащих состоянию субстрата PROGRESSION, появившихся в результате разрастания клеточных множеств, как эффекта активирующих воздействий и совпадающих с ними вершин + состояния PROGRESSION на плоскости III градаций прозрачности ядер. Множество {<0.5x1.0;L=IV>} вершин ** дерева подсостояния PROGRESSION VITAL, пересекло наружную грань C1С20C1 малого пентаэдра, и аналогично множеству клеток ** {<0.4=x>;L=IV} состояния PROMOTION ACTIVE, занимает положение вне малого характеристического пентаэдра на плоскости четвертой градации прозрачности ядер, внутри большого пентаэдра. Множество вершин *** {<0.5x0.8,ILII>}, активируемых агрессивными подмножествами клеток * состояния PROGRESSION, также находится вне объема пентаэдра, в объеме единичного куба на плоскости II или даже I градации прозрачности ядер.
- с. Так как грани пентаэдра, сечения его внутреннего пространства и проекция основания образуют разделяющие поверхности относительно структуры графа, то это означает, что структура графа отражает ситуационную информацию на вершинах, соответствующую классификации образующих граф элементов по состояниям субстрата. Изменение состояния субстрата отражается в наклон и характер треугольных пирамид и многогранника в целом, а направление его изменений отражается направленностью дуг и "свода" графа. Пентаэдр — отражение информационных исполнительных технологий на клеточных множествах.
- d. Объем, покрываемый малой характеристической пирамидой, отделяет множество подмножеств всех отображений текущих изменений от их подмножества VITAL - клеток фатального состояния. Кроме того, на структуре графа это, отделенное множество, разделяется на два множества подмножеств, одно из которых * содержит агрессивные клетки, реализующие активацию процесса роста из состояния PROMOTION ACTIVE, а другое + и ** отражают следствие, результативность этой активации. При этом активация процесса множествами клеток + состояния PROGRESSION, как порожденная, идет в направлении от большей градации оптической плотности к меньшей, т.e. от верхнего шарового слоя к нижнему. Это позволяет говорить о том, что порождение процесса активацией клетками состояния + реализует компоненту разрастания ***, в то время как активация процесса множеством клеток * из состояния PROMOTION ACTIVE порождает компоненту роста. Можно также высказать предположение, что ветвь множества подмножеств клеток ** IV градации оптической плотности отражается в топологическом пространстве накоплением множества клеток, покрываемых шаром локального гомеостазиса, а подмножеств активирующих клеток состояния PROGRESSION и активируемых ими клеток отражают динамику перетоков.
Фокус фазового пространства со всей системой координат также развернулся на 90º и отражается в точку <1.0,IV> на единичной сфере, вне характеристической пирамиды — большого пентаэдра, с координатой 3,33(3)G на плоскости IV градации прозрачности ядер. В нем, как и ранее, отражается множество озлокачествленных клеток, принадлежащих состоянию PROGRESSION VITAL - голых ядер. Полученный многогранник может рассматриваться как полностью определенная характеристическая пирамида канонического состояния размножающегося клеточного множества. Граф, полученный в пространстве характеристической пирамиды, есть полностью определенное отображение в объеме единичного шара, решетки # исходного описания на gl-сагиттальном представлении роста. Аналогичные отображения могут быть получены и для иных фазовых пространственных решеток.
Таким образом, характеристическая пирамида, обладая свойствами разделения клеточных множеств по ролевой функции и компонентам роста, позволяет отразить и численно характеризовать систему межклеточных отношений агрессивности в виде направленного пространственного графа. Вершины графа на мощностях множеств клеток, образующих ситуации, отображают ситуативные отношения между ними одновременно и в терминах текущих состояний и на мере Гомпертца.
Множество межклеточных отношений агрессии отражается на множестве геометрических параметров характеристической пирамиды, образующей ситуативное пространство, в виде классов (категорий) групповых отношений — порождения, развития и озлокачествления. Эти отношения оказались связанными между собой фундаментальным отношением на константах состояний, образовав инварианты процесса. Пространственный граф установил в пространстве характеристической пирамиды отношение соответствия между качественным описанием состояния субстрата и его количественными атрибутами.
Соответствия качественного описания и численных атрибутов в локальном гомеостазисе.
Малая характеристическая пирамида позволяет произвести оценку отношений активации и агрессии. Все исходные данные для расчета численных характеристик на ней, определены в прямоугольной системе координат на мере озлокачествления G. Расстояния на пирамиде определялись на мере Гомпертца, как значения, характеризующие взаимное положение двух конкретных точек в пространстве. Переменные, как и ранее, исчислялись в нормированных значениях. Площади и объемы, покрываемые различными поверхностями, также определены на мере Гомпертца, все отношения имеют соответствующие размерности, отношения констант состояния безразмерны. Это соответствует условию сопоставления на безразмерных значениях констант озлокачествления, и позволяет проводить сравнительный, по относительным величинам, анализ показателей. Приведем некоторые данные.
Координаты центров гомеостазических шаров и параметры их топографии: гомеостазиса — <0.05,II,1.0>; локального гомеостазиса — <1.00,III,1.0>; расстояние между центрами гомеостазических шаров — 3.266 (G), радиус единичного шара фазового пространства равен 3.333(3) (G). Отношение между радиусом единичного шара фазового пространства и межцентровым расстоянием гомеостазических шаров оказалось равным 1.02 и с точностью до погрешностей обработки исходных данных в 2%, соответствует значению раковой константы NCCa для "раковой жемчужины". Это на мере G отражает максимальный нормированный рост, равный 3.33(3)G, достигаемый на древесных порядках и отображенный в фазовое пространство единичного шара. Высота h=-0.50, сегмента шара локального гомеостазиса, с формально приписанным отрицательным значением показателя озлокачествления G, как и положение отсекаемого сегмента в IV квадранте единичного шара, отражают пространство молодых клеток состояния NORMAL решетки исходного описания, как не вступивших в отношения агрессии с множеством клеток баланса эквивалентности и не покрываемых, вследствие этого, единичным кубом. Его значение 0.50G, соответствует нормальному состоянию невозбужденных клеток.
Отношения на элементах архитектуры характеристической пирамиды позволили определить нормированные (по maxG) значения роста G, и "обращенный рост", как обратную ему величину 1/G. Оказалось, что значениям обращенного роста, отраженного в пространстве пирамиды, соответствуют константы состояния. Это свидетельствует, что константы состояния отображаются в топологическое пространство непосредственно. К их числу относятся дисплазионная константа (отношение между радиусами шаров C1C2 и его проекциями), раковая константа (положение центра шара локального гомеостазиса 0C2), а также обращенный рост (положение проекции центра шара локального гомеостазиса С2 на нижней поверхности собственной сферы). Его значение оказалось близким к 1.0, с отклонением не более чем 2%, что отражает завершенность процесса и переход от живого к мертвому. Переходы от живой к неживой материи, рассматриваются в получившей развитие, биоэнергоинформатике — ветви теории торсионного мира, мира кручения. Как мы видели, процессы на информационных технологиях локального гомеостазиса, порождая исполнительные технологии, непрерывно закручивают фазовую траекторию в сторону фокуса. Это приводит сумму технологий озлокачествления в отношения с торсионным миром.
Отношение объема VС1 шара гомеостазиса к объему VС2 шара локального гомеостазиса соответствует значению дисплазионной константы и равно 0.295+-1.37%. Это означает, что обратное соотношение этих объемов соответствует величине радиуса единичного шара — максимальному росту для канонического состояния, выраженного в единицах Гомпертца и равного 3.33(3) G. Отсюда следует вывод, важнейший для задачи компьютеризации профилактической диагностики злокачественных опухолей:
- расстояние между гомеостазическими шарами, обратно пропорционально значению дисплазионной константы, а сама дисплазионная константа есть прогностическое отображение достижимости канонического состояния биологического субстрата.
Отношения агрессивности, порожденная агрессивность активирующих клеток, инварианты.
Значения линейных величин, определенные на характеристической пирамиде, представляют возможности проследить некоторые линейные, площадные и объемные отношения на архитектуре топологического пространства. Вот некоторые из объемных отношений:
- а. Отношение агрессивности, близкое к дисплазионной константе, равное 0.296, определяется фактором активирующих межклеточных взаимодействий клеток множества подмножеств NCR-сагиттальной фазовой решетки с ее (L-1)-ортогональным дополнением;
- в. То же отношение для in-сагиттальной фазовой решетки с ее (L-1)-ортогональным дополнением, равно 0.219 и достаточно близко к константе перехода эпителия спокойного состояния в пластическое;
- с. Относительная агрессивность, значение которой близко к значению раковой константы, характеризуется величиной 0.738. Эта величина отражает отношение активирующих межклеточных взаимодействий клеток in- и NCR-сагиттальных фазовых решеток. Можно предположить, что клетки, озлокачествленные в результате этого взаимодействия, есть клетки разрастания области PROGRESSION VITAL. Если это так, то справедливо утверждение, что клетки, обладающие ядерно-цитоплазматическим индексом близким к раковому, порождены не в процессе роста опухоли, но в процессе ее разрастания, т.е. вторичны. На дереве роста они образуют крону;
- d. Коэффициент порожденной агрессивности, равный 0.401. Значение этого коэффициента близко к значению константы, соответствующей переходу эпителия из состояния пластичности, в состояние гиперпластичности. Он отражает отношение вторичной активирующей интерактивности — порожденной межклеточным взаимодействием множества подмножеств клеток, ранее озлокачествленных в результате активирующей деятельности клеток (L-1)-ортогонального дополнения (NCR)-сагиттальной фазовой решетки и (NCR/in)-пересечения фазовых решеток. Можно полагать, что клетки, озлокачествленные в результате порожденной агрессивности, также есть клетки разрастания;
- e. Коэффициент порожденной агрессивности, аналогичный рассмотренному выше, но относительно in,- и (NCR/in)-пересечения фазовых решеток. Его значение равно 0.297 и близко к значению дисплазионной константы.
Таким образом можно заключить, что характеристическая пирамида, в отличие от характеристической плоскости, разделяет составляющие биологического субстрата не просто по состояниям, отраженным в значения параметров озлокачествления, а по проявляемому ими характеру групповых межклеточных взаимодействий — порождения, развития и озлокачествления, т.е. по агрессивности. Приведенные численные показатели характеризуют систему этих отношений. Эти же соотношения справедливы и относительно большой пирамиды, ибо они отражают линейные отношения на шарах. Рассмотрение приведенных соотношений. позволяет сделать следующий важный вывод:
- константы состояния — ядерно-цитоплазматическая NCI, дисплазионная NCD, раковая NCCa, определенные как моды соответствующих распределений для различных состояний биологического субстрата, а также гомеостазическая константа NCG переключения, отображают активирующие свойства множеств клеток в процессе трехкомпонентного роста, как результата групповых межклеточных взаимодействий, проявляющихся при пересечениях множеств клеток на фазовых пространственных решетках.
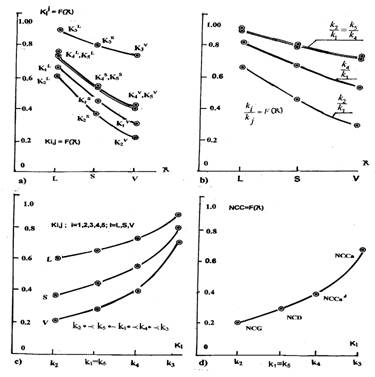 Этот вывод подтверждается зависимостями, приведенными на рисунке 4. Они отражают отношения совершенного строгого порядка на множестве констант состояния, каждой из которых соответствует предмет имени — денотат. В качестве такого предмета имени выступает некоторый, распределенный в пространстве биологического субстрата клеточный объект, объединенный межклеточными отношениями на сетях входа, в виде совокупности клеток "вычлененной", отделенной от множества их подмножеств, и давшей имя той или иной константе состояния. Отношения порядка на множестве констант состояния определены как на отношениях а) и b) "прямой", так и на отношениях c) и e) относительной и порожденной агрессивности, которые образуют категории отношений агрессивности. Это отражается численными значениями соотношений значений коэффициентов порождения (рис.4а), развития (рис.4б) и озлокачествления (рис.4с). Кривые на этих рисунках отражают упорядочение категорий плоскостных L, площадных S и объемных V отношений агрессивности и коэффициентов агрессивности на этих категориях. Из их рассмотрения (рис.4d), можно сделать вывод, что на множестве констант состояния формируется система упорядочения функционально-ролевых отношений агрессии, отношений их следования, а также структурных отношений на множестве коэффициентов вторичной агрессии, порожденных отношениями a), с) и d).
Этот вывод подтверждается зависимостями, приведенными на рисунке 4. Они отражают отношения совершенного строгого порядка на множестве констант состояния, каждой из которых соответствует предмет имени — денотат. В качестве такого предмета имени выступает некоторый, распределенный в пространстве биологического субстрата клеточный объект, объединенный межклеточными отношениями на сетях входа, в виде совокупности клеток "вычлененной", отделенной от множества их подмножеств, и давшей имя той или иной константе состояния. Отношения порядка на множестве констант состояния определены как на отношениях а) и b) "прямой", так и на отношениях c) и e) относительной и порожденной агрессивности, которые образуют категории отношений агрессивности. Это отражается численными значениями соотношений значений коэффициентов порождения (рис.4а), развития (рис.4б) и озлокачествления (рис.4с). Кривые на этих рисунках отражают упорядочение категорий плоскостных L, площадных S и объемных V отношений агрессивности и коэффициентов агрессивности на этих категориях. Из их рассмотрения (рис.4d), можно сделать вывод, что на множестве констант состояния формируется система упорядочения функционально-ролевых отношений агрессии, отношений их следования, а также структурных отношений на множестве коэффициентов вторичной агрессии, порожденных отношениями a), с) и d).
Упорядочение функционально-ролевых отношений образуется путем объединения линейных L отношений порождения, площадных S отношений развития и объемных V отношений озлокачествления. Соотношения коэффициентов порождения, развития и озлокачествления показывают, что линейные и объемные отношения роста связаны кубической зависимостью, отражающей групповые межклеточные отношения агрессивности. Это означает, что значение дисплазионной константы пропорционально кубу коэффициента порождения (L-1)/NCR-агрессивности, значение гомеостазической константы пропорционально кубу (L-1/in)-агрессивности, а раковая константа достаточно приближена к кубу коэффициента порожденной, т.е. вторичной агрессивности взаимодействия множеств клеток (NCR//(NCR/in) -пространственных фазовых решеток. Значение коэффициентов порожденной агрессивности, соответствующее значению дисплазионной константы, в свою очередь равной кубу коэффициента порождения этого вида агрессивности, свидетельствует о наличии параллельно взаимодействующих и равномощных по эффективности множеств клеток, образующих соответствующие пересечения и имеющих различную ролевую направленность. Пространственные пересечения, формируясь клеточными множествами различных сагиттальных направлений роста, содержат объединения и пересечения клеточных множеств, обладающих различными ролевыми функциями.
Система упорядочения констант состояний и их функционально-ролевых отношений, отражает общую упорядоченность функциональных клеточных взаимодействий, реализующих целевую функцию системы локального гомеостазиса, стабилизирующую трехмерный рост.
Так как определение констант проводилось для множества различных локализаций и разновидностей опухолей и подтвердилось на каждой из персоналий, а устойчивость их значений верифицировалось для различных мощностей случайных выборок из генеральной совокупности, с учетом отношений строгого порядка на их множестве, можно говорить о свойстве инвариантности ядерно-цитоплазматических констант. В качестве главного инварианта на топологии карциногенеза выступает межцентровое расстояние С1С2 гомеостазических шаров, численно определенное как 3.266(G) и с погрешностью равной ~2%, не превышающей погрешности обработки клиническихданных, приближенно к радиусу единичного шара фазового пространства. Положение центра шара гомеостазиса и его объем остаются фиксированными как в процессе нормальной жизнедеятельности, так и в процессе размножения клеточных множеств и играют роль инвариантов системы гомеостазиса.
Положение центра шара локального гомеостазиса устанавливается и также остается фиксированным с момента возникновения хотя бы единичных клеток, обладающих раковым отношением, но длина его радиуса варьируется в зависимости от направлений роста и устанавливается в результате их суммарного взаимодействия. Поэтому положение в фазовом пространстве центров гомеостазических шаров, как и их межцентровое расстояние, также может рассматриваться в качестве инварианта системы локального гомеостазиса. Регулярность изменения констант состояния (рис.4d) повторяет характер потери устойчивости проницаемости ядерной мембраны как фактора, обуславливающего переход клеток биологического субстрата в неспокойное, и в последующем — в пластическое и гиперпластическое состояния.
Таким образом, на графе агрессии выявилось множество упорядоченных последовательностей отображений процессов активации, реализуемых на множествах межклеточных отношений биологического субстрата. Полученные закономерности, соответствующие им численные показатели и общие свойства процесса, отраженные в характер и параметры фазовой траектории, изображающей достижение цели управления, находятся в отношении соответствия с множеством отображений отношений агрессии. Это подтверждает высказанные ранее предположения о существовании программного инструментария целевой функции локального гомеостазиса, определяющего инвариантность констант состояния и реализующего, тем самым, фатальную "надежность" функционирования системы стабилизации трехкомпонентного роста.
Функционирование такого инструментально-программного аппарата отображается константами состояния NCC={NCG,NCD,NCCa',NCCa} - инвариантами локального гомеостазиса, образующими систему упорядочения на функционально-ролевых отношениях межклеточных взаимодействий. Эти инварианты отражают состояния биологического субстрата, соответствующие отношениям порождения и перехода от накопленных количественных к существенным качественным изменениям.
Система упорядочения констант состояния, отражая на архитектуре топологии карциногенеза последовательную многофункциональность клеточных множества и мультитехнологичность процессов озлокачествления, находит свою рефлексию на множестве собственных функциональных свойств. Не исключено, что информационная значимость констант, образованная закручиванием фазовой траектории, имеет жизненную важность и отражает взаимосвязи информационно-энергетических характеристик торсионных полей клеточного мира, связанных с информационной насыщенностью, энергетикой процесса озлокачествления и витальностью клеточных множеств кроны древесного роста, в частности с пороговой витальностью, разделяющей живое от неживого.
Яков Гельфандбейн
(c) 1011494 CIPO
Популярное
Страны Евросоюза вновь наращивают импорт удобрений из России. Под разговоры о том, какая это "вредная" продукция и пора поставить ей заслон.



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.