Система локального гомеостазиса как раковый гомеостат
 Системные свойства локального гомеостазиса
Системные свойства локального гомеостазиса
Известный специалист в области математической биологии Н.Рашевски писал, что "...Несмотря на различия, мы находим поразительное относительное подобие между всеми организмами... В общей биологии мы не имеем универсальных констант, таких, какие мы в большом количестве находим в физике. Однако мы находим определенные инвариантные системы отношений между различными свойствами, общими для всех организмов..."
Говоря об инвариантных системах, Рашевски имел в виду системы так называемых "биоинвариантов", биологических свойств организмов, характеризующих организм как целое, подчеркивая их различие по своей физико-химической конституции. Мы, описывая феноменологию частного, — общих механизмов локального гомеостазиса различных органов, установили феномен порождения констант состояния как систему инвариантов организма, относящихся в отношениях с системой биоинвариантов. Аналогичное суждение мы высказали при рассмотрении констант состояния эпителиальных полей подопытных животных.
Инвариантность отражает свойства и отношения в системе. Это понятие эквивалентно понятию симметрии, определяющему статистическое равновесие и отражающему баланс, устанавливающий неизменность свойства порождаемости озлокачествленных клеток. Инвариантность констант состояния в системе локального гомеостазиса порождается преобразованиями симметрии.
Инвариантность существует при соблюдении трех условий: а) когда существует объект преобразования, б) когда существует система отсчета для оценки этих преобразований и в) когда известны собственно преобразования, изменяющие ситуацию на объекте, определяющие возникновение и сохранение инвариантов.
Первые два условия определены: объект преобразования существует в виде клеточного множества, образующего систему локального гомеостазиса. Система отсчета и мера оценки преобразований биологического субстрата, а также мера озлокачествления и константы состояния в этой системе квалифицированы как инварианты динамических отношений по фактическим данным.
Проведем анализ третьего условия — существования и сохранения механизма преобразований, приводящих к формированию устойчивого множества инвариантов как системы, отражающей функцию механизмов локального гомеостазиса. Такой анализ должен отразить механизм динамики смены клеточных ситуаций, отображенный в свойство инвариантности констант состояния.
Системные средства формирования и сохранения инвариантов.
а). Самосохранение и динамичность.
Отношения на клеточных множествах отражают сохранение многообразий ролевых функций на всех этапах развития процесса. Понятие инвариантности выражает самосохраняемость этих отношений и их свойств в ходе преобразований — перетоков и транспорта клеточных множеств. Транспортные ситуации на клеточных множествах направлены на создание инвариантов в новом качестве и на сохранение их функций в последующих перетоках. Обладая свойствами динамичности, сохранение направлено на совершенствование целевой функции локального гомеостазиса и совершенствование способа ее осуществления.
Отображение динамичности сохранения выражается в строгой упорядоченности порождения и передачи преемственности свойств констант состояния. Самосохранение реализуется собственной динамичностью. Принцип сохранения, обладая свойствами динамичности механизма исполнения, превратился в принцип самосохранения развития, эволюции ролевой функции. Принцип эволюции реализуется ретрансляцией функций клеток множеств-денотатов констант состояния баланса эквивалентности в множества-денотаты состояний развития и озлокачествления.
Самосохраняемость — особая форма устойчивости сохранять свое основное качество как следствие внутренних реакций, возникающих в системе в ответ на внешние относительно нее, в том числе, направленные воздействия. Потеря самосохраняемости системой гомеостазиса приводит к гомеостазическому переключению. Система локального гомеостазиса, благодаря установившемуся, после гомеостазического перехода внутреннему равновесию, вступила в новую систему отношений — симметрии ее структуры и возникшей функции. Это отражается достижением однородности ее структурных характеристик. Рассмотрение таких характеристик, позволяет сделать вывод об отражении ими внутреннего равновесия системы локального гомеостазиса, как соответствия ее частей друг другу. Такое равновесие можно сравнить с равновесием термодинамических систем, которое характеризуется макроскопическими параметрами всех ее частей и максимумом энтропии, как функции термодинамического состояния системы в целом. Любая термодинамическая система со временем достигает своего равновесия. Система локального гомеостазиса достигает внутреннего равновесия формированием множества-универсума клеток баланса эквивалентности и первичной сети входа на нем. Сохранение этого равновесия достигается развитием этих сетей. При этом энтропия, как мера неопределенности распределения состояний субстрата — вероятности существования системы в изменяемом качестве, достигает некоторого максимума.
б). Целевая ориентация и эквифинальность.
Самосохранение системы локального гомеостазиса — сущность собственной целевой ориентации. Любая биологическая система, в том числе и система антагонистичная гомеостазису (крайне нежелательная для человека), стремится к "...конечной экологической цели — выживанию" (С.Бир). Ее целевая ориентация, следуя Биру, есть сохранение функционального инварианта локального гомеостазиса, образовавшего клеточную популяцию — вид сообщества. Конечная цель — системообразующий фактор, направленный на формирование и сохранение системно-структурных свойств этой популяции, как элементарной единицы эволюции, способной реагировать на изменения среды.
Движение системы к упорядочению смены состояний различными путями, приводящими к достижению цели, называют эквифинальностью. Биологические системы приходят к одному и тому же конечному состоянию различными путями. В системе локального гомеостазиса такое движение осуществляется по различным сагиттальным направлениям роста. Финальное состояние образует концевой инвариант системы локального гомеостазиса, как форму "гомеостатического" управления в виде пространства, покрываемого сферой локального гомеостазиса объемом в 14,187 G3 единиц Гомпертца.
Свойства самосохранения локального гомеостазиса выражаются в терминах константности. Константность придает процессу направленность и определенность. Пределы качества управления — константные величины, определяемые множествами клеток-денотатов констант состояния. Но определенность есть не только четкое отражение и определение присущих процессу трансформации признаков, но и отрицание признаков, противоположных для данного явления. "Omnis determinatio est negatio" (Spinosa). Каждое явление, наряду с изменениями, сохраняет какие-то основные черты, выступающие как равные самим себе, как те же самые, тождественные, инвариантные. Знание инвариантов в конкретной системе обеспечивается системами отсчета, мерой на них и знанием преобразований, переводящих объект управления из одного состояния в другое. Инварианты, писал Макс Борн "...суть величины, которые имеют одно и то же значение для любой системы отсчета и поэтому независимы от преобразований".
в). Самоорганизация и самовоспроизведение.
Имеются все основания полагать, что для систем клеточного гомеостазиса специфическим является программированность сохранения способа функционирования с предварительным выбором управляющих программ. Алгоритмы этих программ и реализуют условия инвариантности. Обязательным свойством алгоритмов биологических систем, является однозначность результата. Программами служат генетические инструкции, определяющие развитие и поведение систем клеточного множества.
Высшая форма самосохранения — самоорганизация, поддержание уровня организации системы и повышение степени ее организованности. Проявление самоорганизации системы локального гомеостазиса мы уже характеризовали, отразив ее информационно-логические связи и определив ее согласованность с ресурсами мощностей клеточных множеств, вовлеченных в процесс. С одной стороны их исчерпание определяет канонизацию процесса, с другой — достаточность для обеспечения требуемой динамики.
Не исключено, что самоорганизация системы локального гомеостазиса реализуется самовоспроизведением. Процедуры самовоспроизведения могут осуществлять механизмы самосохраняемости, но не просто путем ретрансляции функций клеток множеств-денотатов констант состояния в множества-денотаты других состояний, а путем воспроизведения этих множеств, например на множествах молодых народившихся клеток. Процесс самовоспроизведения при этом осуществляется двумя раздельными операциями — копирования инструкций, содержащих описание воспроизводимого элемента и конструирование этого элемента по заданной инструкции. В системе локального гомеостазиса этому может соответствовать копирование генетической информации, заключенной в молекулах ДНК и передачи ее воспроизводимым элементам. Способность систем поддерживать в определенных пределах характеристические, индикаторные переменные, сформулирована в законе "необходимого разнообразия" Эшби.
Согласно Эшби "...только разнообразие может уничтожить разнообразие". Именно закон необходимого разнообразия позволил установить меру озлокачествления Гомпертца, а сохранение определенных инвариантов на ней, отражает координированность и достижение цели управления осуществляемыми алгоритмами. Система локального гомеостазиса изменяет разнообразия управлений, варьируя мощность множества возбуждаемых клеток так, чтобы распределение частостей показателей состояния, трансформируясь на перетоках, сохраняло свое модальное значение вблизи последовательного ряда значений инвариантов, а значит и количественной меры управления.
Внешние воздействия не порождают обращения состояний системы локального гомеостазиса и это отражает принцип сохранения разнообразия в ней. Этот принцип аналогичен закону сохранения энергии в управляемой системе и именно это делает возбужденный процесс активации необратимым ни при каких внешних воздействиях, направленных на управление им извне: основная функция локального гомеостазиса состоит в самосохранении и эволюции, направленной на совершенствование этой функции.
Статистическая сущность локального гомеостазиса обеспечивает разнообразие связей между его различными функциональными составляющими путем выбора и формирования множества элементов клеточных множеств, участвующих в процессе. Как писал Н.Винер "Связи внутри организации не должны быть абсолютно устойчивыми, чтобы строгая определенность одних ее частей не исключала возможности изменения каких то других. Эти изменения, различные в различных случаях, неизбежно носят статистический характер".
Статистическая природа роста клеточных множеств, связана с иерархическим принципом структурирования его разрастания. Иерархия структуры подчинена отношению следования, следующему из отношения порождения. Она четко просматривается на архитектуре топологии клеточного множества, на которой отображается функциональная иерархия агрессии, как процесса реализации целевой функции локального гомеостазиса. Ярко выраженная динамическая устойчивость системы локального гомеостазиса обусловлена в первую очередь инвариантностью его основных характеристик — констант состояния, как детерминированных характеристик случайного процесса. Это придает системе локального гомеостазиса явно выраженные качества гомеостата.
г). Локальный гомеостазис и гомеостатическая система.
Системы гомеостазиса поддерживают постоянство существенных переменных клеточного сообщества, в локальном гомеостазисе — транспортного потока. Понятие гомеостазиса было развито физиологами К.Бернаром и В. Кэнноном, широко рассматривалось И.М.Сеченовым и И.П.Павловым. Оно связано с понятием ультраустойчивости, адаптивности и стремлением биологических систем к саморегуляции, которая немыслима без организации обратных связей. Ролевая функция гомеостазиса реализуется поиском адекватности вариантов и структур физиологических актов клеточного сообщества условиям внешней среды.
Английский нейрофизиолог У. Росс Эшби разработал электромагнитное устройство, моделирующее свойство гомеостазиса и назвал его "гомеостатом". Это устройство, не делает ничего, кроме непрерывного движения к равновесию. Всего возможно 254=390.625 различных состояний гомеостата Эшби, часть из которых статически устойчива, а часть неустойчива. Если система находится в неустойчивом состоянии, происходит переключение. Переключения происходят до тех пор, пока система не найдет одно из устойчивых состояний вблизи среднего положения. Целеустремленное поведение гомеостата, при котором он находит устойчивое состояние при самых различных возмущениях, Эшби и назвал свойством ультраустойчивости. После Эшби были построены гомеостаты с гораздо большим количеством состояний, в том числе на компьютерах различной мощности и с разным быстродействием. Гомеостат Эшби не имеет памяти, потеряв устойчивое состояние при изменении условий, он возвращается в устойчивое состояние только случайно, в процессе нового поиска.
Для гомеостата Эшби вероятность принятия устойчивого состояния и энтропия являются инвариантами. Если в нем исключить из поиска малоперспективные состояния или реализовать первоочередное опробование перспективных состояний, гомеостат делается способным к обучению, и тем самым к улучшению процесса поиска устойчивых состояний. Если алгоритм поиска первоначально предполагает использование успешных режимов, время случайного поиска сокращается. Беспорядочный случайный поиск в нем нерационален.
Ранее мы предположили, что упреждающая трассировка первоначальных сетей изменения состояний клеточных множеств путем прокладки первичных инициальных сетей входа, одновременно с подготовкой переключающего множества клеток, есть средство предстартовой организации транспортного процесса. Можно предположить, что такая организация создается маркировкой транспортных путей метками множества баланса эквивалентности и формируется как наиболее эффективно решающая задачу ультраустойчивого гомеостатического управления.
Не исключено, что задача гомеостатического управления как задача сохранения, реализуется именно этим множеством, как одна из его ролевых функций. Существование в локальном гомеостазисе гомеостатического управления отражается фиксацией в клинике свойства сохраняемости и хорошо известным свойством его сопротивляемости к воздействиям внешней среды.
Анализ клинических данных и полученных в настоящей работе результатов, дает основание полагать, что система локального гомеостазиса, как механизм естественной технологии, реализует целевую функцию, продуктом которой является непрерывность стабильного сохранения процесса стабилизации неравновесного клеточного роста. Эта система имеет необходимые (и достаточные) признаки естественного гомеостата.
д) Мультиустойчивость.
Система локального гомеостазиса на распределенных множествах клеток-денотатов состояний гомеостатична: она отражает все признаки ультраустойчивости в каждом из последовательно сменяемых текущих состояний. Это означает, что структурно она может состоять из "сборки" ультраустойчивых и независимых друг от друга в определенные промежутки времени, гомеостатических подсистем на сетях входа, обладающих высокой приспособляемостью к параметрам внешней среды и сопротивляемостью межклеточным возмущающим воздействиям, возникающим на этих сетях. Сборка таких подсистем при их параллельно-последовательном функционировании в пространстве субстрата, образует систему, обладающую свойством мультиустойчивости. Работа каждой из них может происходить автономно, вне зависимости от других структурно-функциональных компонент, определяемых различием задач, выполняемых каждым отдельным функциональным подмножеством, устойчивость которого управляется на принципах гомеостата.
Так как система локального гомеостазиса стабилизирует неравновесное состояние роста клеточного множества, это означает, что на заданном уровне поддерживается не отдельный параметр состояния, а протяженный во времени процесс заданного изменения этого параметра, т.е. траектория. Так как траектория изменения состояния, отображаемая следом изображающей точки в фазовом пространстве, отражает стабилизированный и распределенный в пространстве субстрататранспортный поток, то гомеостатическое управление сохраняемостью образует наиболее сложную форму стабильности, которую называют "гоморезом". Это означает, что в системе локального гомеостазиса имеет место стабилизация не каких либо, хотя бы и существенных переменных, а функционального назначения клеточных подмножеств. С этой точки зрения локальный гомеостазис — суть функциональный гомеостат. Понятие гомеостата, при этом, понимается как специфическая программная подсистема.
Следуя Россу Эшби, систему гомеостазиса нужно отнести к классу ультраустойчивых, а систему локального гомеостазиса — к классу мультиустойчивых систем. Сохранение устойчивого неравновесного управления ростом даже при активных репрессивных воздействиях, например средствами химиотерапии, свидетельствует о наличии признаков именно такой системы. Это и порождает мысль о том, что система локального гомеостазиса подстраивается к внешним, относительно этой системы воздействиям, не просто путем всесторонней адаптации к ним, а посредством приспособления, как минимум одной из своих подсистем, в частности — целевого управления, к определенным параметрам этих воздействий и это приспособление реализуется с помощью сборки гомеостатов. Определение характера, особенностей таких воздействий и поиск путей адаптации к ним, могут реализовываться на основе биологической компьютерной системы распознавания, играющей роль подсистемы сохранения, быть может аналогичной той, которая позволяет некоторым живым существам противостоять различного рода химическим средствам борьбы с ними.
Этому в частности не противоречит и тот известный факт, что клеточные множества у некоторых животных, быть может не обладая средствами эффективного гомеостатического приспособления, не всегда могут сопротивляться действию ряда противораковых химиотерапевтических препаратов и системы локального гомеостазиса этих животных не могут быть отнесены к классу ультраустойчивых, тем более мультиустойчивых систем. В то же время, локальный гомеостазис клеточных множеств различных органов человека находит средства приспособления и противостояния. Скорее всего, это связано с организацией "сборок" специфических клеточных гомеостатов с последующей их эволюцией в соответствии с общими принципами эволюции видов.
Гомеостатические механизмы в биологических системах реализуют стабилизацию жизненно важных функций и так обстоит дело в системе гомеостазиса. Система локального гомеостазиса, в результате гомеостазического перехода, меняет свою целевую функцию на антагонистическую, переключаясь с ультраустойчивого управления равновесным состоянием в гомеостазисе, на мультиустойчивость стабилизации неравновесного роста в локальном гомеостазисе, приобретая полный арсенал системных средства формирования и сохранения инвариантов.
Система гомеостазиса может быть отнесена к классу ультраустойчивых систем, система локального гомеостазиса в результате гомеостазического переключения, приобретает новое отличительное качество. Ее можно рассматривать как естественно-технологический "раковый" гомеостат, в котором гомеостатичность, как функция сохранения собственного равновесия и гомеостазичность, как функция сохранения собственного предназначения, находятся в отношении кооперации.
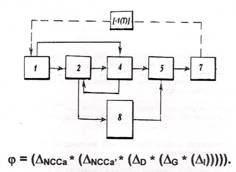
К сожалению, в рамах интернет-статьи невозможно воспроизвести даже малую часть выкладок и рассуждений относительно гипотетических механизмов сохранения тех свойств ракового гомеостазиса, которые обеспечивают достижение финального состояния, образующего концевой инвариант системы. Тем не менее, мы приводим гипотетический логический алгоритм коррекции сохранения (механизм ракового гомеостата), на котором 1,2,4,5,7 — отношения на ситуациях (внутриситуационных перетоках клеточных множеств сохранения); 1-2, 2-4, 4-5, 5-7 — отношения на межситуационных перетоках — транспорте клеточных множеств; 1-4, 4-2, 2-8, 8-5 — отношения управления сохранением многообразий; [-1(Т)] - генеральная обратная связь, 8 — распределенное клеточное множество механизма сохранения. Оцифровка соответствует обозначениям граф-октавы.
Обобщенное управление "фи", сохранением многообразий (функций) размножающегося клеточного множества во времени эволюционного процесса, записывается в виде цепочки воздействий "*" операторов изменений "дельта" I-составляющих управления на внутриситуационных перетоках и в транспорте множеств клеток, обладающих ролевой функций сохранения. Это управление имеет рекурсивный характер, отраженный в возвратную последовательность с большой глубиной (36 возвратов) рекурсии. В каноническом виде оно приведено на рисунке. Из него видно, что гомеостатическая трехкомпонентная (а точнее — четырехкомпонентная) составляющая сохранения, образуемая управлением блока сохранения 8, проявляется на всем множестве шагов рекурсии, охватывающем множество "дельта"-изменений всех состояний, порождаемых клеточным множеством баланса эквивалентности. Схема показывает также, что раковый гомеостат стремится сохранить ролевую функцию активации и порождения с момента возникновения первично порожденных множеством клеток баланса эквивалентности клеток сохранения 1 до момента канонизации процесса. Изменения эволюционных составляющих управлений, записаны здесь с учетом внутренних связей и они носят характер микроэволюционных изменений. Заметим, что это уравнение, несмотря на его внешнюю простоту, настолько сложно, что даже при знании вида образующих его операторов и их характеристик, оно не доступно для решения ни по быстродействию, ни по объему памяти, ни одной из современных ЭВМ. Однако некоторые предположения на нем, делать можно. В частности можно утверждать, что нарушение (разрыв) именно этой рекурсивной цепочки сохранения, создает возможность срыва свойства эквифинальности и прерывания движения системы к финальному состоянию. Меньшей глубине рекурсии соответствует большая вероятность успеха.
Обобщенное управление, как показывают процедуры его построения, базируется на свойствах инвариантов состояния, отражающих их формирование. Инварианты состояния, находятся в отношениях золотого сечения, которое проявляется многократно на многих свойствах клеточных множеств, в частности в бигомеостазическом пространстве, разделяя его на подпространства гомеостазиса и антигомеостазиса.
Американский математик Марк Барр предложил называть отношение золотого сечения, числом "фи" - первой буквой в имени великого Фидия, который часто использовал его в своих скульптурах. Кеплер говорил, что "Геометрия владеет двумя сокровищами: одно из них — теорема Пифагора, другое — деление отрезка в крайнем и среднем отношении. Первое можно назвать мерой золота, второе же больше напоминает драгоценный камень". Именно это отношение определяет двойную оптимальность в системе локального гомеостазиса, и подчеркивая это, мы обозначили обобщенное управление эволюционными процессами в нем, именем "фи" золотого сечения. Отношение золотого сечения проявилось также на отношениях L- и V-витальностей "тонкого" мира. Оно отразило двойную недостаточность логических обоснований этой теории на абстрактном отношении информативности и энергетичности, применительно к процессам перехода клеточных множеств из состояния "живое" в состояние "неживое".
Систематизируем, в самом общем виде, выводы о свойствах системы локального гомеостазиса как ракового гомеостата.
- Условия существования инвариантности на множестве элементов системы локального гомеостазиса, соблюдены. Система локального гомеостазиса, как объект сохранения, определена в качестве естественно-технологической системы патологии, удовлетворяющей постулатам и соглашениям общей теории систем с системой отсчета преобразований, определенной на естественной мере озлокачествления.
Преобразования, изменяющие ситуацию на клеточном объекте, определяющие возникновение и сохранение инвариантов и обуславливающие качество мультиустойчивости локального гомеостазиса, определены как механизм его сохранения, формирующий функциональный "раковый" гомеостат.
- Система гомеостазиса может быть отнесена к классу ультраустойчивых систем, система локального гомеостазиса в результате гомеостазического переключения приобретает новое отличительное качество. Как продукт естественной технологии, эта система реализует единственную функцию — непрерывного стабильного сохранения процесса стабилизации неравновесного роста.
Систему локального гомеостазиса можно рассматривать как естественно-технологический раковый гомеостат, в котором гомеостатичность, как функция самосохранения собственного равновесия и гомеостазичность, как функция сохранения собственного предназначения, находятся в отношении кооперации.
- Самосохранение системы локального гомеостазиса есть сущность собственной целевой ориентации — сохранения клеточных множеств денотатов функциональных инвариантов, породивших разрастающуюся клеточную общность как объект экологии. Конечная экологическая цель — системообразующий фактор, направленный на формирование и сохранение системно-структурных свойств денотатов как элементарной единицы эволюции, способной реагировать на изменения среды.
Механизм самосохраняемости реализуется ретрансляцией функций клеток множеств-денотатов констант состояния баланса эквивалентности в множества-денотаты состояний развития и озлокачествления.
- Инварианты ракового гомеостата отражаются как феномен мультиустойчивости, формируемый, контролируемый и управляемый механизмом сохранения путем трансляции, в порядке их упорядочения, преемственности функций клеточных множеств — денотатов инвариантов и переноса их подстраиваемых значений средствами транспорта клеточных множеств сохранения.
Передача преемственности ролевой функции и перенос подстраиваемых значений инвариантов в каждом отдельном акте возникновения и развития опухоли образует отдельный акт самосовершенствования сохранения.
- Естественно-технологический механизм генерации системы, обладающей свойствами гомеостата, осуществляется целенаправленным управлением сохранения. Это управление реализует адресацию путей трассировки сети управлений к элементам клеточных множеств так, чтобы показатели их состояния оказались подчиненными заданному закону распределения с модальными значениями, определяющими текущие инварианты.
Ролевая функция множества клеток системы сохранения направлена на поддержание заданного значения инвариантов состояния средствами межклеточных взаимодействий, что делает инварианты состояния управляемыми, а фокус фазового пространства абсолютным, обеспечивающим эквифинальность ракового гомеостата.
- Система локального гомеостазиса, как многоканальная мультиустойчивая система стабилизации неравновесного роста, образуется переменным во времени набором автономных гомеостатов, каждый из которых формируется на сети входа одной из целевых клеточных подсистем — денотатов инвариантов. Функционируя независимо, они выполняют задачи статистического сохранения каждого клеточного множества — денотатаинвариантов, регулируя внутриклеточные процессы так, чтобы модальные значения сохраняемых распределений локализовались в пределах, определяющих инвариантность.
Раковый гомеостат функционирует как многоканальный вероятностный регулятор, сохраняющий устойчивость констант состояния активации, порождения и симметрии, как основного условия эквифинальности, соблюдение которого приводит к сохранению раковой константы.
- Как система стабилизации неравновесного роста, раковый гомеостат поддерживает на заданном уровне протяженный во времени процесс наперед заданного изменения состояния — траекторию. Траектория изменения состояния, в виде следа изображающей точки в фазовом пространстве, есть отражение изменений стабилизируемого транспортного потока. Стабилизация транспортных потоков в биологических системах характеризуется сложной формой стабильности — гоморезом.
Раковый гомеостат осуществляет стабилизацию не просто существенных переменных состояния. Он реализует это путем стабилизации свойств собственных отношений относительно функционального назначения определяющих его текущее состояние, собственных клеточных подмножеств. Раковый гомеостат эволюционирует как функциональный гомеостат гомореза.
- Не исключено, что самоорганизация и эволюционное развитие функционального ракового гомеостата реализуются самовоспроизведением, механизм которого осуществляет не просто ретрансляцию функций клеток множеств-денотатов инвариантов, но воспроизводит эти множества как минимум двумя раздельными операциями — генерации и копирования инструкций, содержащих описание корректируемого элемента и конструирования элемента по заданной инструкции.
В эволюционирующей системе локального гомеостазиса этому может соответствовать управление копированием генетической информации, заключенной в молекулах ДНК клеток-денотатов инвариантов и передача ее информационного наполнения воспроизводимым элементам.
- Система локального гомеостазиса, обладая приобретенным качеством функционального ракового гомеостата, поддерживается механизмом цикличного сохранения, воспроизводящего множество его единичных актов эволюции, последовательность которых образует собственные циклы вековой эволюции. Отражение этого механизма может быть получено с помощью граф-октавы в виде многоцикличной смены ситуаций, отношения на которых образуют циклы эволюционирующего качества сохранения.
Негативный феномен эволюции сохранения отражается естественно-технологическим механизмом формирования биологического функционального гомеостата вековым алгоритмическим инструментарием, как специфической целенаправленной системы самосохранения многообразий функции.
- Система локального гомеостазиса формируется как самоорганизующаяся, на свойствах отношения "клеточного золотого сечения" функциональных клеточных множеств баланса эквивалентности, клеточных инвариантов, разделяющей плоскости бигомеостазического пространства и отражается на этих же свойствах графа-октавы. Это отношение отражает условие самоорганизации, образования ракового гомеостата и сохранения локального гомеостазиса, как отношения количественных характеристик размножающихся клеточных множеств противоборствующих клеточных антагонизмов.
Отношение золотого сечения, отражает количественные соотношения между антагонистическими системами гомеостазиса и антигомеостазиса как между противоположностями одного и того же клеточного объекта. Оно является инвариантом — мерой достижения устойчивого (для нас нежелательного) компромисса, отражая общую для всей действительности реально существующего мира, меру компромисса как условия, определяющего смену качества, перехода от живого к неживому. Именно это обуславливает единство структуры отражения разнообразных явлений реального мира моделями в виде граф-октавы на множествах разнообразных отношений, присущих моделируемым объектам.
- Разрушение свойства мультиустойчивости системы локального гомеостазиса, как функционального ракового гомеостата, определяет пути естественного внешнего вмешательства в механизмы его функционирования с целью нарушения реализуемых им функций сохранения инвариантов, а следовательно и свойств эквифинальности.
Нарушение функции сохранения инвариантов в каждом отдельном акте сохранения путем разрушения коммуникативных взаимодействий клеточных множеств-денотатов состояний и, тем самым отношений порождения и развития, образует естественный путь прерывания транспортных потоков клеточных множеств и борьбы с грозным заболеванием.
Яков Гельфандбейн
Популярное
Пугачева выразила свое восхищение выступлением Савельевой, что стало для нее мощной мотивацией в творческом пути.



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.