Древесная архитектура и мера озлокачествления клеточных множеств
 Вероятностные оценки признаков озлокачествления отразили стремление клеточных множеств биологического субстрата к упорядочению на всех этапах своего существования. Оценки на естественно — технологической мере подтвердили такое упорядочение. Они отразили его отношениями совершенного строгого порядка внутренней замкнутости множества клеток нормального состояния на различных этапах метаморфозы, переключающимися на отношение максимального совершенного порядка множества клеток злокачественного роста. Образуется иная функционально направленная внутренняя замкнутость, в пределах которой, на новом составе и новой структуре множеств, устанавливается новая категория отношения порядка.
Вероятностные оценки признаков озлокачествления отразили стремление клеточных множеств биологического субстрата к упорядочению на всех этапах своего существования. Оценки на естественно — технологической мере подтвердили такое упорядочение. Они отразили его отношениями совершенного строгого порядка внутренней замкнутости множества клеток нормального состояния на различных этапах метаморфозы, переключающимися на отношение максимального совершенного порядка множества клеток злокачественного роста. Образуется иная функционально направленная внутренняя замкнутость, в пределах которой, на новом составе и новой структуре множеств, устанавливается новая категория отношения порядка.
Концепция "хаотичности" роста в процессе трансформации биологического субстрата не находит своего подтверждения, порядки на клеточных множествах биологического субстрата управляемы.
Гомеостазический переход, осуществляющий переключение отношений порядка, отражает ситуационный конфликт двух противоречивых и противоборствующих механизмов. Один из них — потери, в момент переключения, упорядоченности на уровне внутриклеточных отношений совершенного строгого порядка подмножеством клеток эпителия инициального состояния. Другой — одновременного установления упорядоченности на уровне межклеточных групповых взаимодействий новой внутренней замкнутости — максимального совершенного порядка на подмножествах активируемых клеток. Общее условие реализации такого перехода требует наличия "15% клеток состояния initiation и соблюдения некоторых дополнительных условий снижения прозрачности ядер в ближайших окрестностях значения 0.30 ядерно-цитоплазматического индекса.
Мы отмечали существование специфического соотношения мощностей клеточных множеств в субстрате различных состояний, обладающих значениями ядерно-цитоплазматического индекса, близкими к тем или иным значениям ядерно-цитоплазматических констант. Спокойное состояние, при этом соотношении, отражается эпителиальной константой для 60% клеток. Пластическое состояние клеток, не перешедших в гиперпластическое состояние, отражается дисплазионной константой для 36% клеток. Состояние озлокачествления гиперпластических клеток, отражается раковой константой для множества такой же мощности в 36%, а голым ядрам соответствует 13% клеток канонического состояния. Можно с большой долей уверенности утверждать, что такое соотношение отражает активизацию равномощного множества клеток конечного продукта.
Но такое предположение порождает и другое допущение: равномощность множества активаторов и активированных клеточных множеств, соответствует энергетическому балансу, достигаемому на множестве клеток баланса эквивалентности.
И здесь нельзя обойти вниманием следующий факт: суммарное количество из 36% клеток, обладающих значением ЯЦИ, равным раковой константе, и количества голых ядер в 13% (с погрешностью, до точности исходных данных в 2.5%), соответствует относительному количеству клеток в 47%, образующих подмножество состояния PROMOTION множества клеток баланса эквивалентности. Это отражает баланс энергии, затрачиваемой на активацию по принципу "клетка на клетку".
С другой стороны, суммарное множество клеток нормального состояния этого баланса, и клеток, перешедших в гиперпластическое состояние, с отклонением примерно в 8% (суммарной методической и инструментальной погрешностей), составляет вторую его половину. Но можно полагать, что энергия для жизнеобеспечения уже озлокачествленных клеток этого баланса не требуется, и она перераспределяется на множестве вновь порождаемых клеток. Поэтому, с некоторой долей уверенности можно также предположить, что баланс энергии на множестве клеток баланса эквивалентности, имеет двойной характер: кроме баланса энергии, затрачиваемой на активацию, он обеспечивает энергетический баланс нарождения молодых клеток и их жизнедеятельности, как общего резерва активирующих клеток состояния PROMOTION. Фланговое положение активирующего множества клеток этого состояния, позволяет связать его с явлением ангиогенеза, как источника такой энергии.
Предположение об активизации клетками-активаторами равномощного множества клеток конечного продукта, следующее из полученных наблюдениями соотношений мощностей клеточных множеств в окрестностях значений ядерно-цитоплазматических констант, и вытекающие из него следствия, образуют ядро клеточного феномена "0.3-0.8".
Рассмотрим отражение механизмов клеточного роста в системе локального гомеостазиса на древесных порядках, с позиций исключительной значимости клеточного множества баланса эквивалентности, порождающего феномен "0.3-0.8". Отметим при этом, что принадлежность феномена локального гомеостазиса к категории систем, как объекта, отражается на постулатах и соглашениях общей теории систем, как биологический порядок, клеточная структура на множестве клеток, связанных функцией сохранения агрессивных групповых взаимодействий, направленных на организацию и поддержание упорядоченных отношений стабилизации неравновесного роста. Эта система представляет собой определенную целостность, сохраняющую единство на всем отрезке времени своего существования, причем свойства этой системы определяются не просто суммой свойств ее элементов, но динамизмами множества их взаимодействий. Наиболее наглядно такие взаимодействия отражаются древесными порядками. Построение древесных порядков предполагает учет всего множества возможных сагиттальных представлений роста, и введение некоторых специфических отношений на деревьях.
Отображение функциональных отношений активации групповыми межклеточными взаимодействиями подмножеств, активирующих процесс образования клеток состояния PROMOTION, объединенных в подмножестве баланса эквивалентности (и образующих в динамике роста правостороннее охватывающее множество), а также отражение динамики отношений клеточных множеств иных состояний, требует введения единой системы индикации фаз, уровней и страт состояний и их идентификаторов.
Будем полагать, что сеть коммуникационных связей решетки множеств размножающихся клеток, свойствами селективности выбора направления роста не обладает, а процесс трансформации реализуется как максимально быстротечный, в результате суммирования одновременных приростов активированных клеток по всем образуемым составляющим. Поэтому, для каждого из сагиттальных представлений, сложность порожденного им дерева, можно трактовать, как одну из компонент параллельного суммарного прироста на множестве парциальных составляющих роста.
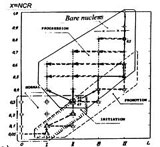
Для создания системы индикации изменений состояния субстрата, а также идентификаторов этих изменений, детализируем дерево роста, в котором множества клеток состояний PROMOTION и PROGRESSION (как объектов активации и подпитки множествами клеточных подмножеств охватывающего состояния PROMOTION), структурно разделены, образуя специфическую древесную архитектуру. Такое дерево показано на рисунке.
В процессе древесного роста, вслед за уровнем нормального состояния N0, появляется вершина NI инициализации процесса — корень дерева (уровень N1). От корня дерева порождается росток ветви инициации, образующий страту P1,11¸P3,13, — порождения множества клеток баланса эквивалентности. В узле разрастания PG0 этой ветви, на уровне P4, появляются три ствола роста клеточных множеств состояния PROGRESSION и правонаправленная "голая" ветвь роста множества клеток подпитки — состояния PROMOTION, не имеющая ветвей.
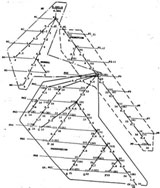 Вершины дерева, отраженные на каждом из пересечений ветви с текущим уровнем роста, образуют фазы состояния. Они оцифрованы значениями сложности, мощности множества клеток прироста и принадлежностью к той или иной градации прозрачности ядер, что идентифицирует их состав и функциональную роль множества клеток, образующих фазу состояния. Уровни идентифицированы именами состояний. Прирост мощности клеточных множеств в пространстве состояния PROGRESSION, образует крону дерева и конечными вершинами отражает редукцию <¬> роста. Знак редукции отражает неиспользованный резерв мощности множества клеток роста на соответствующей вершине. Межуровневые интервалы характеризуют прирост каждой отдельной ветви и оцениваются накопленными значениями веса, помечающими дуги. В качестве весов дуг выбраны значения ядерно-цитоплазматического индекса, характерного для межуровневого интервала.
Вершины дерева, отраженные на каждом из пересечений ветви с текущим уровнем роста, образуют фазы состояния. Они оцифрованы значениями сложности, мощности множества клеток прироста и принадлежностью к той или иной градации прозрачности ядер, что идентифицирует их состав и функциональную роль множества клеток, образующих фазу состояния. Уровни идентифицированы именами состояний. Прирост мощности клеточных множеств в пространстве состояния PROGRESSION, образует крону дерева и конечными вершинами отражает редукцию <¬> роста. Знак редукции отражает неиспользованный резерв мощности множества клеток роста на соответствующей вершине. Межуровневые интервалы характеризуют прирост каждой отдельной ветви и оцениваются накопленными значениями веса, помечающими дуги. В качестве весов дуг выбраны значения ядерно-цитоплазматического индекса, характерного для межуровневого интервала.
Структура дерева, образующая его архитектуру, включает, таким образом, пространство четырех поддеревьев — состояний NORMAL NN гомеостазиса, INITIATION NIN - образования множества клеток-активаторов баланса эквивалентности, PROMOTION NPR - активирующего и подпитывающего процесс и состояния PROGRESSION NPRG - локального гомеостазиса, как состояния активированного новообразования. Эти пространства образуются четырьмя множествами функциональных клеточных подмножеств NN, NIN, NPR и NPRG. Вершине разрастания соответствует уровень роста PRG0=P4, соответствующий узлу сцепления <0.3,II> на характеристической плоскости, порождающему пространство состояния PROGRESSION. Порождающая прогрессивный рост, и четко разграничивающая клеточные множества на функциональные подмножества, ролевая функция множества баланса эквивалентности в узле сцепления <0.3,II>, отражает сущность клеточного феномена "0.3-0.8".
Отображение роста древесными порядками на текущем множестве n уровней, порождается трехкомпонентным ростом мощности клеточных множеств и изменением состояний в функции текущего времени t, отображение которого соответствует отображению изменений. Состояния и их изменения по уровням, именуются идентификаторами уровней. Идентификатор уровня отражает образ состояния, конкатенированный образами состояния фаз, создающих уровень. На дереве роста, как уже отмечалось, текущее время образует параметр уровня.
Свобода выбора сагиттальных представлений, определяющих скорость достижения канонического состояния, создает систему предпочтений, отражающих отношение порядка между парциальными составляющими роста. Предпочтения выявляют те или иные преимущества факторов влияния, определяющих скорость роста, как отражения следа перемещения на решетке множеств при различных сагиттальных представлениях, с учетом принятых ранее договоренностей. Характерное число уровней роста n зависит от вида сагиттального представления. При принятом уровне конкретизации, оно получило два значения и оказалось равным 10 и 13.
Из множества сагиттальных направлений роста, мы выделили специфическое, обуславливающее агрессивный активирующий характер клеток состояния PROMOTION ("глобальная составляющая" роста, см. титульный рисунок). Эта составляющая отражает "скоростной" процесс озлокачествления в общем направлении "наискорейшего спуска" с "затягиванием". При этом учитывалось, что изменения имеют необратимый характер, т.к. клетки, подвергшиеся трансформации не восстанавливаются. Из этого сагиттального направления роста видно, что ядерно-цитоплазматическое отношение клеток и прозрачность их ядер, находятся в отношении порядка, образуя дважды упорядоченные множества параметра состояний <N<i, j>, Þ, ¬>, где < Þ > строгий порядок, а < ¬ > редукция.
Древесная архитектура феномена метаморфозы образуется на множестве текущих древесных порядков персоналий. Множество древесных порядков на множестве сагиттальных представлений, образует модель древесной архитектуры. Не имея возможности получить древесную архитектуру по персоналиям, под понятием "древесная архитектура", будем понимать ее модель на множестве сагиттальных представлений.
В терминах общей теории систем, древесные порядки соответствуют отображению отношений порождения на сетях коммуникационных взаимодействий выхода W и входа Г системы локального гомеостазиса на сети изменения состояния.
Таким образом, древесный порядок отражает динамику стабилизации трехкомпонентного неравновесного роста по каждому из его стволов и ее упорядоченность, эксплицирует трехмерный рост, конкретизирует его как исходное понятие трехмерного фазового пространства.
Удалось установить некоторые формальные отношения, которые с позиций теории множеств показывают, что отображение множества состояний субстрата, как максимального совершенного подмножества, удовлетворяет при любой последовательности идентификаторов попарно непересекающихся совокупностей фаз состояния на своей области определения (уровне дерева), условию равносильности отображения объединения состояний отдельных подмножеств (фаз) суммой отображений фаз их состояний.
Текущее состояние s отражается конкатенацией фаз, а множество состояний S, оказалось счетно-аддитивной функцией на подмножестве текущих состояний. Так как отображение состояний определено на совокупности подмножеств значений обобщенного показателя озлокачествления N такой, что она удовлетворяет определенным условиям объединения любого числа своих элементов, то текущее состояние s определяет меру (в теории множеств — кольцо множеств).
С позиций исключительной значимости клеточного множества баланса эквивалентности, это означает, что текущее состояние s, как мера, на всем множестве S текущих состояний, оценивает текущие отношения эквивалентности и толерантности, определяющие состояния субстрата, ставя в соответствие фиксированным значениям s или приращениям состояний Ds, некоторые эквивалентные единицы, в общем случае не обязательно числа. Это могут быть, например, множества, отраженные на графиках соответствия или кривые, в цифре отображающие изменения состояния субстрата, или их отрезки, идентифицированные именами фазовых переходов, фаз или страт m при фиксированных значениях текущих состояний s. При этом, все сказанное относительно состояния как меры, справедливо и для отображения времени. Интервалы времени Dt изменения состояния, есть мера времени.
Для сравнительного оценивания древесных порядков на мере состояния s, мы использовали определенную систему обозначений, отличающих отображения возможных сагиттальных направлений роста, а в качестве единицы измерения, для сопоставления отображений различных парциальных составляющих процесса, — интервалы Ds. Эти интервалы переменны, в зависимости от изменения межуровневого расстояния изменений состояний, идентифицируемых именами уровней. Это предопределило принципиальную возможность ввести специфическую для процесса озлокачествления, естественно — технологическую численную меру и единицу измерения на ней.
 Сопоставление приращений древесных порядков, полученных на множестве сагиттальных представлений роста, отражающих изменение состояний относительно среднего роста, позволяет сделать ряд выводов относительно активности порождения, как результата реализации функции управления процессом стабилизации роста.
Сопоставление приращений древесных порядков, полученных на множестве сагиттальных представлений роста, отражающих изменение состояний относительно среднего роста, позволяет сделать ряд выводов относительно активности порождения, как результата реализации функции управления процессом стабилизации роста.
Для такого сопоставления мы приняли пять вероятных представлений роста, основным из которых является глобальное представление glS, а также парциальные направления накопления ядерно-цитоплазматического отношения NCRS и снижения прозрачности ядер LS, а также направления CanS, отражающего процесс активации методом "прямого" возбуждения (см. рисунок).
 Сопоставление, в частности, позволило оценить, на отношении следования, как на отношении предпочтения, сравнительное "быстродействие" компонент роста, его влияние на "темп" озлокачествления, а также форсировку процессов активации и достижения канонического состояния. Оказалось, что высота дерева для различных составляющих может отличаться на одну — три фазы. Это следует из того, что в парциальных составляющих с меньшим числом фаз, имеет место более интенсивное сцепление фаз состояния, и это следствие повышенной агрессивности подмножеств активирующих клеток состояния PROMOTION, образующих баланс эквивалентности, по сравнению с клетками подмножеств того же состояния, охватывающих множество фаз подпитки.
Сопоставление, в частности, позволило оценить, на отношении следования, как на отношении предпочтения, сравнительное "быстродействие" компонент роста, его влияние на "темп" озлокачествления, а также форсировку процессов активации и достижения канонического состояния. Оказалось, что высота дерева для различных составляющих может отличаться на одну — три фазы. Это следует из того, что в парциальных составляющих с меньшим числом фаз, имеет место более интенсивное сцепление фаз состояния, и это следствие повышенной агрессивности подмножеств активирующих клеток состояния PROMOTION, образующих баланс эквивалентности, по сравнению с клетками подмножеств того же состояния, охватывающих множество фаз подпитки.
Кроме того, выявлены различия между составляющими роста, отображенного в текущие состояние s для различных сагиттальных представлений. Они свидетельствую о наличии, в биологическом субстрате процессов, быть может, и не главенствующих, но влекущих за собой первичность снижения прозрачности ядер клеток по отношению к ядерно-цитоплазматическому отношению, особенно на конечных стадиях процесса. Эти различия представляются существенными.
Оказалось, что наиболее интенсивный характер процесса трансформации (относительно других его сагиттальных составляющих), отражает парциальная составляющая, отмеченная как glS - "глобальная" (что и обусловило ее название), с подпиткой озлокачествляющихся клеток из фаз охватывающего состояния PROMOTION. Разрастание опухоли при этом представлении, наиболее интенсивно и идет со значительным опережением относительно других направлений разрастания, с ускорением на начальных стадиях озлокачествления. Это подтверждает агрессивный характер поведения множества клеток состояния PROMOTION баланса эквивалентности. В частности, состояние субстрата, соответствующее фазе (уровню) PRG0, достигается при этом направлении разрастания в 5.5 — 8.0 раз быстрее, чем при всех иных парциальных составляющих процесса. Иллюстрация этого факта приводится на рисунке. Именно это определяет опасность раннего образования, хотя бы и одиночных клеток состояния PROMOTION.
Суммарное отображение древесного роста, полученное как нормированное среднее, а также изменение (приращение) состояний, в связи с таким свойством глобального направления сагиттального роста, обладают S-образной формой, что указывает на наличие колебательной составляющей относительно других парциальных составляющих. Оно достаточно близко по своему характеру к составляющей накопления ядерно-цитоплазматического отношения NCRS, вначале опережая его примерно на 0.5 уровня, а затем, на интервале [5-8] уровней, отставая от него, причем это отставание при m=5, составляет почти целый уровень, увеличиваясь до полутора уровней при m=8. Далее это отставание сокращается, по-видимому, за счет форсировок роста. Такой характер отображения суммарного роста свидетельствует о главенствующем влиянии на начальном этапе, когда еще не сформировались агрессивные множества клеток охватывающего состояния PROMOTION, более "инертных" составляющих — снижения прозрачности ядер LS и направления CanS, отражающего процесс активации методом возбуждения, а при превышении четвертого уровня — о резко возрастающей агрессивности глобальной glS составляющей.
Это позволяет предположить, что активность управления процессом стабилизации роста в системе локального гомеостазиса, возрастает постепенно. Она зависит от динамики формирования сетей входа Г и выхода W по всем возможным парциальным направлениям разрастания, по мере накопления мощности множеств фаз состояния, включившихся в межклеточные взаимодействия на формирующейся сети коммутативного обмена управляющей информацией. При этом, процесс ускорения роста имеет инерционный и плавный характер: отображение суммарного роста и все другие его отображения, не имеют скачков в момент гомеостазического перехода.
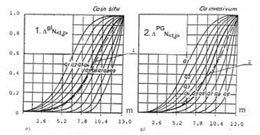 Мера s представляет практическую возможность для сопоставительного численного анализа различных факторов, сопутствующих трассировке и формированию отображений рассмотренного типа. Отображения приращений мощности множеств на глобальном glS сагиттальном направлении роста, относительно общего приращения мощности клеточного множества состояния PROGRESSION, есть функция, отображающая прирост сложности дерева в пространстве состояний.
Мера s представляет практическую возможность для сопоставительного численного анализа различных факторов, сопутствующих трассировке и формированию отображений рассмотренного типа. Отображения приращений мощности множеств на глобальном glS сагиттальном направлении роста, относительно общего приращения мощности клеточного множества состояния PROGRESSION, есть функция, отображающая прирост сложности дерева в пространстве состояний.
В частности, разница между отображением суммарного роста и парциальной составляющей glS, определяет прирост сложности дерева за счет разрастания мощности подмножеств фаз состояния по уровням, в то время как приращение показателя озлокачествления для состояния PROGRESSION, определяет изменение роста дерева по высоте. Эти две кривые имеют интересную особенность: нанесенные на сетку распределения вероятностей озлокачествления, они практически совпадают с распределением, рассчитанным, с достоверностью по вероятности равной единице (жирная линия), определяющим модельный эталон распределения вероятностей для рака.
Это, однако, и не удивительно: по своей природе все отображения на феномене озлокачествления, имеют одно и то же происхождение. Они лишь получены, исходя из различных предпосылок.
Яков Гельфандбейн
(c) 1011494 CIPO
Популярное
Стюардессы поделились советами о том, как успешно справиться с проблемой перевеса ручной клади при посадке на борт самолета.



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.