Порядок в мире клеточных популяций. Граф-диагност
Для биоматематиков
 Ученые солидарны в своем мнении относительно путей исследования мироздания. Знаменитый американский физик – теоретик, лауреат Нобелевской премии Ричард Филлипс Фейнман - один из основателей квантовой электродинамики, писал, что в явлениях природы есть формы и ритмы, недоступные глазу созерцателя, но открытые глазу аналитика.
Ученые солидарны в своем мнении относительно путей исследования мироздания. Знаменитый американский физик – теоретик, лауреат Нобелевской премии Ричард Филлипс Фейнман - один из основателей квантовой электродинамики, писал, что в явлениях природы есть формы и ритмы, недоступные глазу созерцателя, но открытые глазу аналитика.
Одним из таких ритмов является ситуативная клеточная динамика - такой механизм размножения клеточного множества, при котором непрерывно меняются не только размытость отдельных клеток, их подмножеств или множеств, но на сетях межклеточных взаимодействий формируются четко выраженные группировки клеточных подмножеств, также четко выраженного функционального назначения. Эти группировки находятся в отношениях, отраженных изменениями обобщенного параметра озлокачествления.
Профессор кибернетики и психиатрии Иллинойского университета Уильям Росс Эшби говорил, что “…если биология призвана изучить и понять подлинно сложные системы, то этому должны соответствовать применяемые ею методы”. Такие методы реализуются общей теорией систем, позволяющей изучить недоступные глазу наблюдателя ритмы клеточных преобразований.
Обычно статические объекты отражают деревьями. Динамика наглядно отражается древесным ростом или направленным графом. При этом преобразования большого количества клеток, как реакции на первичный акт активации процесса, выявляет не просто динамичность, но ее кинетический характер. Деревья и графы находятся в отношении соответствия, дерево – разновидность графа.
Процессы ситуативной клеточной динамики отражаются, как активируемые, во времени и в дискретном численном пространстве. Они соответствуют причинно-следственным связям функциональных отношений, образуемых множеством групповых взаимодействий. Эти взаимодействия направлены не только на увеличение текущей мощности подмножеств озлокачествляемых клеток, уже дифференцированных по возрастающим значениям аргумента обобщенного показателя озлокачествления, но и на генерацию их новых подмножеств, в частности – из молодых клеток, путем вовлечения их в процесс озлокачествления. Такие отношения наглядно отображаются с помощью графов.
Рассортированные на характеристическом поле канонического состояния, группировки клеточных подмножеств позволили изучить их свойства, определить узлы сцеплений и пересечения, отразить статику завершенного процесса на множестве признаков, определенном на всем множестве смен состояний. Однако особый интерес представляет ситуативная динамика и кинетика процесса трансформации всего множества клеточных подмножеств, участвующих в процессе, включая финитные подмножества тех клеток, которые к моменту канонизации оказались замороженными в своем текущем состоянии.
Это означает, что ситуативная клеточная динамика должна отображать самую смену состояний. Такое отображение должно характеризовать динамику возникновения и формирования комбинаций множеств стартовых подмножеств клеток из состояния нормы в состояние пластичности, из состояния пластичности в состояние гиперпластичности и перетоки из них, как из состояний отправления в состояния прибытия. При этом, должно найти свое отражение появление молодых клеток, а также новых множеств клеток, находящихся в функциональных отношениях. Это осуществляется отличением от понятия “рост” понятия “разрастания”. Если рост обусловлен транспортом клеточных подмножеств, поступающих в новое состояние как во множество прибытия, то разрастание отражает дальнейшее накопление мощности множества клеток за счет процессов, происходящих в пределах каждого из достигнутых состояний. Эта составляющая отражает непрерывность процесса формирования и накопления мощности текущих стартовых множеств на множестве достигнутых состояний.
Под механизмом трансформации одиночной клетки понимается такой процесс ее озлокачествления, который, будучи активирован извне или в результате самоактивации под воздействием сопутствующих внешних факторов, приводит к изменению ее состояния, отраженного в изменение парных значений признаков - аргументов обобщенного параметра озлокачествления. В результате этих изменений клетка включаются в одно из клеточных подмножеств, вовлекаемых в процесс трансформации биологического субстрата.
Под механизмом активации, соответственно, понимается порожденный межклеточными информационными взаимодействиями единичный (или множественный) акт инициации процесса, направленного на трансформацию одиночных клеток или их множества. Механизм активации генерируется управлениями, внешними, относительно каждой отдельной клетки или их множества. Множества актов трансформации и активации обуславливают ситуативность динамических процессов, находящихся в отношениях. Эти отношения отражается графом. Они определяют правила назначения направления его дугам.
Механизмы трансформации и активации позволили выявить связи, отражающие непрерывность преобразования биологического субстрата во времени и пространстве, и представить преобразования на подмоделях двух параллельных подпроцессов, находящихся в причинно-следственном отношении порождения. Одна из них отражает процесс трансформации, в котором направление дуг модели всегда совпадает с направлением роста значений парных признаков, обуславливающих значение обобщенного признака Ni,j озлокачествления, независимо от характера устойчивости процесса, другая отражает процесс его активации, направление которой определяется отношениями клеточных подмножеств, образующих фазы состояний. Это привело к рассмотрению двух логически согласованных актов активации – “возбуждением” и “затягиванием”.
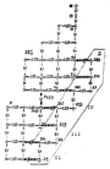 На заглавном рисунке приведено дерево кинетики озлокачествления, построенное на характеристической сетке решетки множеств. Параметром является изменение прозрачности ядер. Стрелки показывают направление перехода каждого элемента прообраза в элемент образа, а представление транспорта множеств с помощью стрелок называют ”сагиттальным”. На характеристической сетке можно выделить некоторое множество сагиттальных направлений, среднее из которых, может считаться оценкой общего направления клеточного транспорта. На каждом из них может быть построен граф. Обобщенный граф, как отображение процесса, показывает, что активация осуществляется благодаря ее внешней подпитке на четырех уровнях состояний, причем замкнутая энергетическая система подпитки реализуется на множестве клеток баланса эквивалентности, перетекающих во множество баланса толерантности. Такой переток отражается узлом инициализации на решетке множеств.
На заглавном рисунке приведено дерево кинетики озлокачествления, построенное на характеристической сетке решетки множеств. Параметром является изменение прозрачности ядер. Стрелки показывают направление перехода каждого элемента прообраза в элемент образа, а представление транспорта множеств с помощью стрелок называют ”сагиттальным”. На характеристической сетке можно выделить некоторое множество сагиттальных направлений, среднее из которых, может считаться оценкой общего направления клеточного транспорта. На каждом из них может быть построен граф. Обобщенный граф, как отображение процесса, показывает, что активация осуществляется благодаря ее внешней подпитке на четырех уровнях состояний, причем замкнутая энергетическая система подпитки реализуется на множестве клеток баланса эквивалентности, перетекающих во множество баланса толерантности. Такой переток отражается узлом инициализации на решетке множеств.
Обобщенный граф отражает структуры взаимодействий клеточных множеств на всех этапах клеточных преобразований, с учетом вовлечения в них порождаемых молодых клеток, образующих резерв множества клеток активации. Это означает образование, уже на начальной стадии процесса, замкнутой управляющей системы, обеспечивающей не только регуляцию процесса активации при его порождении, но и ее устойчивое функционирование, защищенное от внешних воздействий. Устойчивость и непрерывность функционирования этого механизма энергетически дублируется: последовательным порождением узлов сцепления активирующих клеток с одной стороны, и одновременным накоплением мощности подмножеств составляющих эти узлы клеток, с другой. Это – первый, инициальный этап трансформации биологического субстрата IN.
Следовательно, важнейшим для профилактики, является не допустить возникновения и формирования клеточного множества баланса эквивалентности (узел инициализации между четвертым и пятым уровнями состояний) и перетока клеток в множество баланса толерантности (пятый этап множества состояний). До возникновения такого баланса и отправления стартового множества транспорта из него, процесс обратим. Подсчет мощностей соответствующих множеств (осуществим инструментально), позволяет установить этот момент четко.
Охватывающий, “фланговый“ характер расположения развивающегося множества активирующих узлов на графе и динамика их роста, отражают функциональную роль клеточных множеств активации, как системы сохранения устойчивости - обеспечения надежности функционирования всего механизма озлокачествления. Его появление свидетельствует о формировании механизма сохранения мультиустойчивости процесса (гомеостат). Не исключено, что такое фланговое отображение положения множества клеток - активаторов на графе, отражает и топографическую специфику его распределения в пространстве субстрата. Образование открытого, со стороны инициального состояния, охватывающего множества клеток-активаторов, образует второй подготовительный и, одновременно, интенсифицирующий этап клеточного роста. Это – этап развития РR. На этом этапе важнейшим является подавление нарождающихся клеток-активаторов “охватывающего“ множества. Такое подавление, однако, возможно только специфическими методами, адекватными методам активации.
Третий, исполнительный этап трансформации субстрата, реализуется на трех нижних уровнях состояния прогрессирования PRG, а ее четвертый, завершающий этап, отражается как устойчивый переходный процесс исполнения целевой функции множества клеток-активаторов, завершивших свое функциональное предназначение. Подавление клеток-активаторов этих уровней весьма проблематично, хотя бы и с применением методов, аналогичных для предыдущего этапа.
Таким образом, процесс озлокачествления реализуется активацией клеточного множества из узла инициации с одной стороны, его поддержанием агрессивным охватывающим множеством клеток-активаторов с другой, в условиях постоянной подпитки агрессии множествами нарожденных молодых клеток. Этот вывод – принципиален.
Отношения на графе позволили установить стремление клеточных множеств к упорядочению на всех этапах метаморфозы. Однако это упорядочение различно на различных ее этапах: отношения совершенного строгого порядка внутренней замкнутости множества клеток нормального состояния, скачкообразно переключаются на отношение максимального совершенного порядка множества клеток размножающегося множества. Отражение этого переключения – инвариант “левизны-правизны“ (равный 0.5 ± 0.025), поддается инструментальному контролю.
Образуется иная функционально направленная внутренняя замкнутость, в пределах которой, на новом составе и новой структуре множеств, устанавливается новая категория отношения порядка. Концепция "хаотичности" роста в процессе трансформации биологического субстрата не находит своего подтверждения. Подавление процесса озлокачествления – суть разрыв отношений упорядочения на клеточных множествах. Разрыв этих отношений возможен только средствами его порождения. Этот вывод также принципиален.
Переход из состояния гомеостазиса в антагонистическое ему состояние антигомеостазиса, отражает ситуационный конфликт двух противоречивых и противоборствующих механизмов. Один из них - потери, в момент переключения, упорядоченности на уровне внутриклеточных отношений совершенного строгого порядка подмножеством клеток эпителия инициального состояния, другой - установления при этом упорядоченности на уровне межклеточных групповых взаимодействий новой внутренней замкнутости - максимального совершенного порядка на подмножествах озлокачествляющихся клеток. Общее условие реализации такого (гомеостазического) перехода, требует наличия »13% клеток инициального состояния и удовлетворения некоторых специфических условий снижения прозрачности ядер, на интервале 0.28 £ x £ 0.32 значений ядерно-цитоплазматической (дисплазионной) константы (равной 0.3 ± 0.02).
Таким образом, множеству текущих состояний биологического субстрата и множеству их смен, соответствует механизмы двух состояний (видов) гомеостазиса: внутренней замкнутости множества клеток равновесного состояния - (H) гомеостазис и внутренней замкнутости множества клеток неравновесного состояния, локализованный на локально разрастающихся патологических образованиях - локальный (LH) гомеостазис. Переключающая функция – суть феномен замещения (H) гомеостазиса локальным (LH) гомеостазисом.
Важно, что переход субстрата из состояния гомеостазиса в состояние локального гомеостазиса, отражает феномен гомеостазического перехода, как следствие изменения отношений порядка, дочерних относительно отношений порождения. Метаморфоза биологического субстрата – суть управляемый и упорядоченный во времени и в пространстве устойчивый процесс стабилизации неравновесного роста. Управляемость и упорядоченность процесса отображаются в максимальной совершенной упорядоченности подмножеств клеток, отражающей отношения порядка функциональных взаимодействий порождения. Сохранение, на множествах максимальных совершенных порядков трансформирующихся структур, является определяющим в механизмах озлокачествления.
Формально состояние биологического субстрата следует понимать как отражения, в пространства состояний и времени, степени его поражения на трехкомпонентных древесных порядках. Эти порядки отображают динамизмы роста - разрастание, прирост и направление. Динамизмы роста, в свою очередь, отображаются в трехмерное пространство изменения состояний, параметров этих изменений и времени, образуя единое формальное пространство. В общей теории систем такое пространство (множество) соответствует категории "фазового" пространства.
Фазовое пространство образуется на соответствующей тройке множеств, находящихся в отношениях порождения и упорядочения, отраженными на криволинейной поверхности, “натянутой" на решетку # графика соответствия сагиттального представления групповых взаимодействий. Полученная "оболочка" покрывает упорядоченное множество текущих изменений степени поражения биологического субстрата во времени.
Формальное представление феномена локального гомеостазиса в виде тройки дважды упорядоченных множеств - степени поражения, состояния и времени, определенных на решетке # множества клеток исходного описания для фиксированного cагиттального представления роста, позволяет отразить в нем динамизмы кинетических процессов. Такое отображение феномена локального гомеостазиса в фазовое пространство, означает строгое разрешение специфической размытой ситуации на множестве нечетких элементов с нечеткими границами.
Установившееся понятие опухоли, справедливо лишь частично: ему соответствует потеря упорядоченности на уровне внутриклеточных отношений. Однако в момент гомеостазического перехода, этот вид упорядоченности скачкообразно замещается упорядочением межклеточных отношений функциональных взаимодействий. Гомеостазический переход не просто отражает возникновение процесса озлокачествления, но множеством клеток баланса эквивалентности генерирует его, а множеством клеток баланса толерантности, поддерживает и питает. Внутриклеточные отношения первичны, межклеточные отношения множеств дифференцированных клеток (групповые взаимодействия) - вторичны.
В терминах общей теории систем опухоль - суть гомеостазическое переключение феномена гомеостазиса в феномен локального гомеостазиса изолированным переключающим подмножеством клеток критической мощности. С позиций теории регулирования, такое переключение означает замену механизма линейной регуляции динамических внутриклеточных процессов, механизмами программного задания и стабилизации, относительно этого задания, неравновесного состояния роста клеточных множеств путем организации и мультиустойчивого поддержания специфических кинетических процессов активации.
Прерывание процесса преобразования биологического субстрата, требует предупреждения гомеостазического переключения, либо разрушения, после реализованного переключения, механизма складывающихся межклеточных отношений на уровне групповых взаимодействий.
Яков Гельфандбейн
© 1011494 CIPO
Популярное
Существует обширное разнообразие пород собак в мире.



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.